
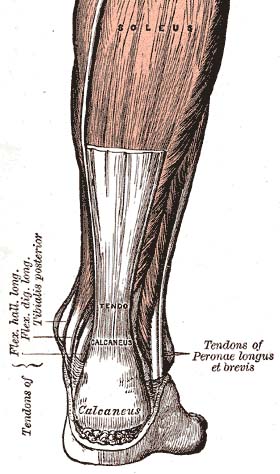

tendo
D013710
H3.03.00.0.00020
Anatomická terminologie
Šlacha nebo šlacha je pevný pruh vláknité pojivové tkáně, který spojuje sval s kostí a je schopen odolávat tahu.
Šlachy jsou podobné vazům; obě jsou tvořeny kolagenem. Vazy spojují jednu kost s druhou, zatímco šlachy spojují sval s kostí.
Struktura
Histologicky se šlachy skládají z hustého pravidelného vaziva. Hlavní buněčnou složkou šlach jsou specializované fibroblasty zvané tenocyty. Tenocyty syntetizují extracelulární matrix šlach, která je hojně tvořena hustě uspořádanými kolagenními vlákny. Kolagenová vlákna jsou navzájem rovnoběžná a uspořádaná do fasciklů. Jednotlivé fascikly jsou spojeny endotendinem, což je jemné volné vazivo obsahující tenká kolagenní vlákna a elastická vlákna. Skupiny fasciklů jsou ohraničeny epitenonem, což je obal z husté nepravidelné pojivové tkáně. Celá šlacha je uzavřena fascií. Prostor mezi fascií a šlachovou tkání je vyplněn paratenonem, tukovou areolovou tkání. Normální zdravé šlachy jsou ukotveny ke kosti pomocí Sharpeyových vláken.
Extracelulární matrix
Suchá hmota normálních šlach, která tvoří 30-45 % jejich celkové hmotnosti, je tvořena:
- 60-85 % kolagenu
- 60-80 % kolagenu I
- 0-10 % kolagenu III
- 2 % kolagenu IV
- malé množství kolagenů V, VI a dalších
- 15-40 % nekolagenních složek extracelulární matrix, včetně:
- 3 % proteinu oligomerní matrix chrupavky,
- 1-2 % elastinu,
- 1-5 % proteoglykanů,
- 0.2% anorganických složek, jako je měď, mangan a vápník.
Přestože kolagen I tvoří většinu kolagenu ve šlachách, je přítomno mnoho menších kolagenů, které hrají zásadní roli ve správném vývoji a funkci šlach. Patří mezi ně kolagen typu II v chrupavčitých zónách, kolagen typu III v retikulinových vláknech cévních stěn, kolagen typu IX, kolagen typu IV v bazálních membránách kapilár, kolagen typu V v cévních stěnách a kolagen typu X v mineralizované fibrochrupavce v blízkosti rozhraní s kostí.
Ultrastruktura a syntéza kolagenu
Kolagenová vlákna se spojují do makroagregátů. Po sekreci z buňky, štěpené prokolagenními N- a C-proteázami, se molekuly tropokolagenu spontánně sestavují do nerozpustných fibril. Molekula kolagenu je asi 300 nm dlouhá a 1-2 nm široká a průměr vytvořených fibril se může pohybovat v rozmezí 50-500 nm. Ve šlachách se pak fibrily dále sestavují do fasciklů, které mají délku asi 10 mm a průměr 50-300 μm, a nakonec do šlachového vlákna o průměru 100-500 μm.
Kolagen ve šlachách drží pohromadě pomocí proteoglykanů (sloučenina tvořená bílkovinou navázanou na glykosaminoglykanové skupiny, přítomná zejména v pojivové tkáni) složek včetně dekorinu a ve stlačených oblastech šlachy také agrekanů, které jsou schopny se navázat na kolagenové fibrily ve specifických místech. Proteoglykany jsou s kolagenními fibrilami provázány – jejich postranní glykosaminoglykanové (GAG) řetězce mají mnohočetné interakce s povrchem fibril – což ukazuje, že proteoglykany jsou strukturálně důležité pro propojení fibril. Hlavními složkami GAG ve šlachách jsou dermatan sulfát a chondroitin sulfát, které se spojují s kolagenem a podílejí se na procesu sestavování fibril během vývoje šlach. Předpokládá se, že dermatan sulfát je zodpovědný za vytváření asociací mezi fibrilami, zatímco chondroitin sulfát se více podílí na obsazování objemu mezi fibrilami, aby je udržel oddělené a pomohl odolávat deformaci. Dermatan sulfátové postranní řetězce dekorinu se v roztoku agregují a toto chování může napomáhat při sestavování kolagenových vláken. Když jsou molekuly dekorinu vázány na kolagenovou fibrilu, mohou se jejich dermatan sulfátové řetězce prodloužit a spojit s dalšími dermatan sulfátovými řetězci dekorinu, který je vázán na oddělené fibrily, a vytvořit tak interfibrilární můstky a nakonec způsobit paralelní uspořádání fibril.
Tenocyty
Tenocyty produkují molekuly kolagenu, které se agregují od konce ke konci a ze strany na stranu a vytvářejí kolagenové fibrily. Svazky fibril jsou uspořádány do vláken, mezi nimiž jsou těsně zabaleny protáhlé tenocyty. Ve šlaše je trojrozměrná síť buněčných procesů spojených s kolagenem. Buňky spolu komunikují prostřednictvím gap junctions a tato signalizace jim dává schopnost detekovat mechanickou zátěž a reagovat na ni.
V endotendonu lze zviditelnit krevní cévy probíhající paralelně s kolagenními vlákny, s občasnými větvícími se příčnými anastomózami.
Předpokládá se, že vnitřní hmota šlachy neobsahuje žádná nervová vlákna, ale epitenon a paratenon obsahují nervová zakončení, zatímco na spojnici mezi šlachou a svalem jsou přítomny Golgiho šlachové orgány.
Délka šlachy se liší u všech hlavních skupin a u jednotlivých osob. Délka šlachy je v praxi rozhodujícím faktorem, pokud jde o skutečnou a potenciální velikost svalu. Například při rovnosti všech ostatních relevantních biologických faktorů bude mít člověk s kratší šlachou a delším bicepsovým svalem větší potenciál svalové hmoty než člověk s delší šlachou a kratším svalem. Úspěšní kulturisté budou mít obecně kratší šlachy. Naopak ve sportech vyžadujících od sportovců vynikající výkony, jako je běh nebo skok, je výhodné mít nadprůměrně dlouhou Achillovu šlachu a kratší lýtkový sval.
Délka šlach je dána genetickou predispozicí a nebylo prokázáno, že by se zvětšovala nebo zmenšovala v závislosti na prostředí, na rozdíl od svalů, které se mohou zkracovat v důsledku úrazu, nerovnováhy v používání a nedostatečné regenerace a protahování. RAT
Funkce

Tradičně byly šlachy považovány za mechanismus, kterým se svaly spojují s kostí, stejně jako svaly samotné, fungující pro přenos sil. Toto spojení umožňuje šlachám pasivně modulovat síly během lokomoce a zajišťovat dodatečnou stabilitu bez aktivní práce. V posledních dvou desetiletích se však mnoho výzkumů zaměřilo na elastické vlastnosti některých šlach a jejich schopnost fungovat jako pružiny. Ne všechny šlachy musí plnit stejnou funkční úlohu, přičemž některé převážně polohují končetiny, například prsty při psaní (polohovací šlachy), a jiné fungují jako pružiny pro zefektivnění lokomoce (šlachy uchovávající energii). Šlachy uchovávající energii mohou uchovávat a obnovovat energii s vysokou účinností. Například během lidského kroku se Achillova šlacha natahuje při dorzální flexi hlezenního kloubu. Během poslední části kroku, kdy se chodidlo plantárně ohýbá (směřuje prsty dolů), se uložená elastická energie uvolňuje. Kromě toho, protože se šlacha natahuje, je sval schopen fungovat s menší nebo dokonce žádnou změnou délky, což umožňuje svalu generovat větší sílu.
Mechanické vlastnosti šlachy závisí na průměru a orientaci kolagenních vláken. Kolagenní vlákna jsou navzájem rovnoběžná a těsně uspořádaná, ale vykazují vlnovitý vzhled způsobený plošným zvlněním neboli krimpy v měřítku několika mikrometrů. Ve šlachách mají kolagenová vlákna určitou flexibilitu díky absenci hydroxyprolinových a prolinových zbytků na specifických místech v sekvenci aminokyselin, což umožňuje vznik dalších konformací, jako jsou ohyby nebo vnitřní smyčky v trojité šroubovici, a vede ke vzniku krimpsů. Krimpy v kolagenových vláknech umožňují šlachám určitou pružnost a zároveň nízkou tuhost v tlaku. Kromě toho, protože šlacha je vícevláknová struktura složená z mnoha částečně nezávislých fibril a fasciklů, nechová se jako jediná tyčinka a tato vlastnost také přispívá k její pružnosti.
Pro mechanické vlastnosti jsou důležité také proteoglykanové složky šlach. Zatímco kolagenní vlákna umožňují šlachám odolávat namáhání v tahu, proteoglykany jim umožňují odolávat namáhání v tlaku. Tyto molekuly jsou velmi hydrofilní, což znamená, že mohou absorbovat velké množství vody, a proto mají vysoký poměr bobtnání. Protože jsou nekovalentně vázány na fibrily, mohou se reverzibilně asociovat a disociovat, takže můstky mezi fibrilami mohou být přerušeny a znovu vytvořeny. Tento proces se může podílet na tom, že se fibrily pod napětím prodlužují a zmenšují svůj průměr. Proteoglykany však mohou mít také vliv na tahové vlastnosti šlachy. Struktura šlachy je ve skutečnosti vláknový kompozitní materiál, který je tvořen řadou hierarchických úrovní. Na každé úrovni hierarchie jsou kolagenní jednotky spojeny buď kolagenními příčnými vazbami, nebo proteoglykany, a vytvářejí tak strukturu vysoce odolnou vůči tahovému zatížení. Bylo prokázáno, že prodloužení a deformace samotných kolagenových vláken jsou mnohem nižší než celkové prodloužení a deformace celé šlachy při stejném množství napětí, což dokazuje, že matrice bohatá na proteoglykany musí také podléhat deformaci a při vysokých deformačních rychlostech dochází k jejímu ztuhnutí. K této deformaci nekolagenní matrice dochází na všech úrovních hierarchie šlach a modulací organizace a struktury této matrice lze dosáhnout různých mechanických vlastností požadovaných pro různé šlachy. Ukázalo se, že šlachy uchovávající energii využívají značné množství posuvu mezi fascikly, aby umožnily požadované vysoké deformační vlastnosti, zatímco polohové šlachy se více spoléhají na posuv mezi kolagenními vlákny a fibrilami. Nedávné údaje však naznačují, že šlachy uchovávající energii mohou obsahovat také fascikly, které jsou zkroucené nebo šroubovicové povahy – uspořádání, které by bylo velmi výhodné pro zajištění chování podobného pružení, které je u těchto šlach požadováno.
Mechanika
Mechanika
Mechanika
Šlachy jsou viskoelastické struktury, což znamená, že vykazují jak elastické, tak viskózní chování. Při natažení vykazují šlachy typické chování „měkké tkáně“. Křivka síla-roztažení nebo křivka napětí-deformace začíná oblastí s velmi nízkou tuhostí, protože se narovnává zvlněná struktura a kolagenní vlákna se vyrovnávají, což naznačuje záporný Poissonův poměr ve vláknech šlachy. Nedávno provedené testy in vivo (pomocí MRI) a ex vivo (pomocí mechanických testů různých šlachových tkání kadaverů) ukázaly, že zdravé šlachy jsou vysoce anizotropní a vykazují záporný Poissonův poměr (auxetický) v některých rovinách při natažení do 2 % podél své délky, tj. v normálním rozsahu pohybu. Za touto oblastí „špičky“ se struktura stává výrazně tužší a má lineární křivku napětí a deformace, dokud nezačne selhávat. Mechanické vlastnosti šlach se značně liší, protože jsou přizpůsobeny funkčním požadavkům šlachy. Šlachy uchovávající energii bývají pružnější nebo méně tuhé, aby mohly snáze uchovávat energii, zatímco tužší polohové šlachy bývají o něco viskoelastičtější a méně pružné, aby mohly zajistit jemnější kontrolu pohybu. Typická šlacha uchovávající energii selže při zhruba 12-15% deformaci a napětí v oblasti 100-150 MPa, ačkoli některé šlachy jsou výrazně roztažitelnější než tato hodnota, například povrchový digitální flexor u koně, který se při cvalu roztáhne o více než 20 %. Polohové šlachy mohou selhat již při deformaci 6-8 %, ale mohou mít moduly v oblasti 700-1000 MPa.
Několik studií prokázalo, že šlachy reagují na změny mechanického zatížení růstovými a remodelačními procesy, podobně jako kosti. Konkrétně studie ukázala, že nepoužívání Achillovy šlachy u potkanů vedlo ke snížení průměrné tloušťky svazků kolagenních vláken tvořících šlachu. U lidí bylo v experimentu, v němž byli lidé vystaveni simulovanému prostředí mikrogravitace, zjištěno, že tuhost šlach se výrazně snížila, a to i v případě, že subjekty musely provádět klidová cvičení. Tyto účinky mají význam v různých oblastech, od léčby pacientů upoutaných na lůžko až po návrh účinnějších cvičení pro astronauty.
Léčení
Šlachy na chodidle jsou velmi složité a komplikované. Proto je proces hojení porušené šlachy dlouhý a bolestivý. Většina lidí, kterým se nedostane lékařské pomoci během prvních 48 hodin od poranění, bude trpět silným otokem, bolestí a pocitem pálení v místě poranění.
Mělo se za to, že šlachy nemohou procházet obratem matrix a že tenocyty nejsou schopny opravy. Od té doby se však ukázalo, že po celý život člověka tenocyty ve šlaše aktivně syntetizují složky matrix, stejně jako enzymy, jako jsou matrix metaloproteinázy (MMP), mohou matrix degradovat. Šlachy jsou schopny se hojit a zotavovat ze zranění v procesu, který je řízen tenocyty a jejich okolní extracelulární matrix.
Tři hlavní fáze hojení šlach jsou zánět, oprava neboli proliferace a remodelace, které lze dále rozdělit na konsolidaci a zrání. Tyto fáze se mohou navzájem překrývat. V první fázi se do místa poranění rekrutují zánětlivé buňky, jako jsou neutrofily, spolu s erytrocyty. Během prvních 24 hodin se rekrutují monocyty a makrofágy a dochází k fagocytóze nekrotických materiálů v místě poranění. Po uvolnění vazoaktivních a chemotaktických faktorů je zahájena angiogeneze a proliferace tenocytů. Tenocyty se poté přesunou do místa poranění a začnou syntetizovat kolagen III. Po několika dnech začíná fáze reparace nebo proliferace. V této fázi se tenocyty podílejí na syntéze velkého množství kolagenu a proteoglykanů v místě poranění a hladiny GAG a vody jsou vysoké. Přibližně po šesti týdnech začíná fáze remodelace. První částí této fáze je konsolidace, která trvá přibližně šest až deset týdnů po úrazu. Během této doby dochází ke snížení syntézy kolagenu a GAG a také ke snížení buněčnosti, protože tkáň se stává více vláknitou v důsledku zvýšené produkce kolagenu I a fibrily se zarovnávají ve směru mechanického namáhání. Poslední fáze zrání nastává po deseti týdnech a během této doby dochází ke zvýšení zesíťování kolagenních vláken, což způsobuje, že tkáň je tužší. Postupně, přibližně během jednoho roku, se tkáň změní z fibrózní na jizvu.
Matrixové metaloproteinázy (MMPs) hrají velmi důležitou roli v degradaci a remodelaci ECM během procesu hojení po poranění šlachy. Některé MMP, včetně MMP-1, MMP-2, MMP-8, MMP-13 a MMP-14, mají kolagenázovou aktivitu, což znamená, že na rozdíl od mnoha jiných enzymů jsou schopny degradovat fibrily kolagenu I. Na rozdíl od jiných enzymů mají kolagenázovou aktivitu. Degradace kolagenních fibril pomocí MMP-1 spolu s přítomností denaturovaného kolagenu jsou faktory, o nichž se předpokládá, že způsobují oslabení ECM šlachy a zvýšení možnosti vzniku další ruptury. V reakci na opakované mechanické zatížení nebo zranění mohou být tenocyty uvolňovány cytokiny, které mohou indukovat uvolňování MMP, což způsobuje degradaci ECM a vede k opakovanému zranění a chronickým tendinopatiím.
Na opravě a regeneraci šlach se podílí řada dalších molekul. Bylo prokázáno, že během hojení šlach je významně zvýšená regulace a aktivita pěti růstových faktorů: inzulinu podobného růstového faktoru 1 (IGF-I), destičkového růstového faktoru (PDGF), vaskulárního endoteliálního růstového faktoru (VEGF), základního fibroblastového růstového faktoru (bFGF) a transformujícího růstového faktoru beta (TGF-β). Všechny tyto růstové faktory mají během procesu hojení různou roli. IGF-1 zvyšuje produkci kolagenu a proteoglykanů v první fázi zánětu a PDGF je rovněž přítomen v časných fázích po poranění a podporuje syntézu dalších růstových faktorů spolu se syntézou DNA a proliferací šlachových buněk. Je známo, že tři izoformy TGF-β (TGF-β1, TGF-β2, TGF-β3) hrají roli při hojení ran a tvorbě jizev. O VEGF je dobře známo, že podporuje angiogenezi a indukuje proliferaci a migraci endoteliálních buněk, a bylo prokázáno, že mRNA VEGF je exprimována v místě poranění šlach spolu s mRNA kolagenu I. Kostní morfogenetické proteiny (BMP) jsou podskupinou nadrodiny TGF-β, které mohou indukovat tvorbu kostí a chrupavek i diferenciaci tkání, a konkrétně u BMP-12 bylo prokázáno, že ovlivňuje tvorbu a diferenciaci šlachové tkáně a podporuje fibrogenezi.
Vliv aktivity na hojení
Na zvířecích modelech byly provedeny rozsáhlé studie zkoumající vliv mechanické zátěže v podobě úrovně aktivity na zranění a hojení šlach. Zatímco protahování může narušit hojení během počáteční zánětlivé fáze, ukázalo se, že řízený pohyb šlach přibližně po týdnu od akutního poranění může pomoci podpořit syntézu kolagenu tenocyty, což vede ke zvýšení pevnosti v tahu a průměru zhojených šlach a menšímu množství srůstů než u šlach, které jsou znehybněné. U chronických poranění šlach bylo rovněž prokázáno, že mechanická zátěž stimuluje proliferaci fibroblastů a syntézu kolagenu spolu s přeskupením kolagenu, což podporuje opravu a remodelaci. Pro další podporu teorie, že pohyb a aktivita napomáhají hojení šlach, bylo prokázáno, že imobilizace šlach po zranění má často negativní vliv na hojení. U králíků se u imobilizovaných kolagenních fasciklů prokázala snížená pevnost v tahu a imobilizace má také za následek nižší množství vody, proteoglykanů a kolagenních příčných vazeb ve šlachách.
Jako příčina reakce tenkocytů na mechanickou sílu bylo navrženo několik mechanotransdukčních mechanismů, které jim umožňují měnit genovou expresi, syntézu proteinů a buněčný fenotyp a nakonec způsobit změny ve struktuře šlachy. Hlavním faktorem je mechanická deformace extracelulární matrix, která může ovlivnit aktinový cytoskelet, a tím ovlivnit tvar, pohyblivost a funkci buněk. Mechanické síly se mohou přenášet prostřednictvím míst fokální adheze, integrinů a buněčných spojů. Změny aktinového cytoskeletu mohou aktivovat integriny, které zprostředkovávají „outside-in“ a „inside-out“ signalizaci mezi buňkou a matrix. Důležité mohou být také G-proteiny, které vyvolávají vnitrobuněčné signální kaskády, a iontové kanály, které se aktivují protažením a umožňují vstup iontů, jako je vápník, sodík nebo draslík, do buňky.
Společnost a kultura
Šindele byly hojně využívány v celé předindustriální éře jako pevné a odolné vlákno. Mezi některá specifická použití patří použití šlachy jako nitě pro šití, připevňování peří k šípům (viz fletch), přivazování ostří nástrojů k násadám atd. V příručkách pro přežití je také doporučována jako materiál, z něhož lze vyrobit pevné provazy na předměty, jako jsou pasti nebo obytné stavby. Tendon musí být upraven specifickým způsobem, aby mohl být pro tyto účely užitečný. Inuité a další obyvatelé cirkumpolárních oblastí využívali šlachu jako jedinou šňůru pro všechny domácí účely z důvodu nedostatku jiných vhodných zdrojů vláken v jejich ekologickém prostředí. Pružné vlastnosti jednotlivých šlach se využívaly také ve složených opakovacích lucích, které si oblíbili stepní kočovníci Eurasie a indiáni. Pružných vlastností šlach využívalo i první dělostřelectvo vrhající kameny.
Šlachy jsou vynikajícím provaznickým materiálem ze tří důvodů: Je mimořádně pevná, obsahuje přírodní lepidla a při schnutí se smršťuje, takže není třeba dělat uzly.
Kulinářské využití
Šlacha (zejména hovězí) se používá jako jídlo v některých asijských kuchyních (často se podává v restauracích yum cha nebo dim sum). Jedním z oblíbených pokrmů je suan bao niu jin, ve kterém se šlacha marinuje v česneku. Někdy se také vyskytuje ve vietnamském nudlovém pokrmu phở.
Klinický význam
Poranění
Šlachy jsou předmětem mnoha typů poranění. Existují různé formy tendinopatií neboli poranění šlach v důsledku nadměrného používání. Tyto typy poranění obvykle vedou k zánětu a degeneraci nebo oslabení šlach, což může nakonec vést k jejich ruptuře. Tendinopatie mohou být způsobeny řadou faktorů týkajících se extracelulární matrix (ECM) šlach a jejich klasifikace je obtížná, protože jejich příznaky a histopatologie jsou často podobné.
První kategorií tendinopatií je paratenonitida, která označuje zánět paratenonu neboli paratendinového listu nacházejícího se mezi šlachou a jejím pouzdrem. Tendinóza označuje nezánětlivé poškození šlachy na buněčné úrovni. Degradace je způsobena poškozením kolagenu, buněk a cévních složek šlachy a je známo, že vede k ruptuře. Pozorování šlach, které prošly spontánní rupturou, ukázala přítomnost kolagenových vláken, která nejsou ve správné paralelní orientaci nebo nejsou rovnoměrně dlouhá či průměrná, spolu se zaoblenými tenocyty, dalšími buněčnými abnormalitami a prorůstáním cév. U jiných forem tendinózy, které nevedly k ruptuře, byla rovněž zjištěna degenerace, dezorientace a ztenčení kolagenních vláken spolu se zvýšením množství glykosaminoglykanů mezi vlákny. Třetím typem je paratenonitida s tendinózou, u níž je přítomna kombinace zánětu paratenonu a degenerace šlachy. Poslední je tendinitida, která označuje degeneraci se zánětem šlachy i narušením cév.
Tendinopatie mohou být způsobeny několika vnitřními faktory včetně věku, tělesné hmotnosti a výživy. Vnější faktory často souvisejí se sportem a zahrnují nadměrné síly nebo zatížení, špatné tréninkové techniky a podmínky prostředí.
Ostatní živočichové

U některých organismů, z nichž vynikají ptáci a ornitischánští dinosauři, mohou části šlach zkostnatět. Při tomto procesu osteocyty pronikají do šlachy a ukládají kost stejně jako v sezamské kosti, například v čéšce. U ptáků dochází k osifikaci šlach především v zadní končetině, zatímco u ornitischánních dinosaurů tvoří osifikované šlachy axiálních svalů mřížku podél neurálních a hemálních trnů na ocase, pravděpodobně kvůli podpoře.
Viz také
| Wikimedia Commons má média související s tématem Šlachy. |
- Aponeuróza
- Chrupavka
- Chordae tendineae
- Seznam svalů lidského těla
- Šlachová pochva
- ^ Dorlands Medical Dictionary, strana 602
- ^ Caldini, E. G.; Caldini, N.; De-Pasquale, V.; Strocchi, R.; Guizzardi, S.; Ruggeri, A.; Montes, G. S. (1990). „Distribution of elastic system fibres in the rat tail tendon and its associated sheaths“ (Distribuce vláken elastického systému ve šlachách potkana a jeho přidružených pochvách). Cells Tissues Organs. 139 (4): 341-348. doi:10.1159/000147022. PMID 1706129.
- ^ Grant, T. M.; Thompson, M. S.; Urban, J.; Yu, J. (2013). „Elastická vlákna jsou široce distribuována ve šlachách a vysoce lokalizována kolem tenocytů“. Journal of Anatomy. 222 (6): 573-579. doi:10.1111/joa.12048. PMC 3666236. PMID 23587025.
- ^ Dorlands Medical Dictionary 2012. strana 1382
- ^ a b c Jozsa, L., and Kannus, P., Human Tendons: Anatomie, fyziologie a patologie. Human Kinetics: Champaign, IL, 1997.
- ^ Lin, T. W.; Cardenas, L.; Soslowsky, L. J. (2004). „Biomechanika poranění a opravy šlach“. Journal of Biomechanics. 37 (6): 865-877. doi:10.1016/j.j.jbiomech.2003.11.005. PMID 15111074.
- ^ Kjær, Michael (duben 2004). „Role extracelulární matrix v adaptaci šlach a kosterních svalů na mechanické zatížení“. Physiological Reviews. 84 (2): 649-698. doi:10.1152/physrev.00031.2003. ISSN 0031-9333. PMID 15044685.
- ^ Taye, Nandaraj; Karoulias, Stylianos Z.; Hubmacher, Dirk (leden 2020). „Těch „dalších“ 15-40 %: The Role of Non-Collagenous Extracellular Matrix Proteins and Minor Collagens in Tendon“ (Úloha nekolagenních proteinů extracelulární matrix a minoritních kolagenů v šlachách). Journal of Orthopaedic Research. 38 (1): 23-35. doi:10.1002/jor.24440. ISSN 0736-0266. PMC 6917864. PMID 31410892.
- ^ Fukuta, S.; Oyama, M.; Kavalkovich, K.; Fu, F. H.; Niyibizi, C. (1998). „Identifikace kolagenů typů II, IX a X v místě inzerce hovězí achillovy šlachy“. Matrix Biology. 17 (1): 65-73. doi:10.1016/S0945-053X(98)90125-1. PMID 9628253.
- ^ Fratzl, P. (2009). „Celulóza a kolagen: od vláken ke tkáním“. Current Opinion in Colloid & Interface Science (Aktuální názory v oblasti vědy o rozhraní). 8 (1): 32-39. doi:10.1016/S1359-0294(03)00011-6.
- ^ Zhang, G. E., Y.; Chervoneva, I.; Robinson, P. S.; Beason, D. P.; Carine, E. T.; Soslowsky, L. J.; Iozzo, R. V.; Birk, D. E. (2006). „Decorin reguluje sestavování kolagenových vláken a získávání biomechanických vlastností během vývoje šlach“. Journal of Cellular Biochemistry. 98 (6): 1436-1449. doi:10.1002/jcb.20776. PMID 16518859. S2CID 39384363. CS1 maint: multiple names: authors list (link)
- ^ Raspanti, M.; Congiu, T.; Guizzardi, S. (2002). „Strukturní aspekty extracelulární matrix šlachy : studie atomárních sil a skenovací elektronové mikroskopie“. Archives of Histology and Cytology. 65 (1): 37-43. doi:10.1679/aohc.65.37. PMID 12002609.
- ^ Scott, J. E. O., C. R.; Hughes, E. W. (1981). „Proteoglykan-kolagenové uspořádání ve vyvíjející se šlaše ocasu potkana. An electron microscopical and biochemical investigation“. Biochemical Journal. 195 (3): 573-581. doi:10.1042/bj1950573. PMC 1162928. PMID 6459082. CS1 maint: multiple names: authors list (link)
- ^ Scott, J. E. (2003). „Elasticita v extracelulární matrix ‚tvarových modulů‘ šlach, chrupavek atd. A sliding proteoglycan-filament model“. Journal of Physiology. 553 (2): 335-343. doi:10.1113/jphysiol.2003.050179. PMC 2343561. PMID 12923209.
- ^ McNeilly, C. M.; Banes, A. J.; Benjamin, M.; Ralphs, J. R. (1996). „Tendon cells in vivo form a three dimensional network of cell processes linked by gap junctions“. Journal of Anatomy. 189 (Pt 3): 593-600. PMC 1167702. PMID 8982835.
- ^ „Having a short Achilles tendon may be an athlete’s Achilles heel“ (Mít krátkou Achillovu šlachu může být Achillovou patou sportovce). Retrieved 2007-10-26.
- ^ Young, Michael. „A Review on Postural Realignment and its Muscular and Neural Components“ (PDF).
- ^ Thorpe C.T., Birch H.L., Clegg P.D., Screen H.R.C. (2013). Úloha nekolagenní matrix ve funkci šlach. Int J ExpPathol. 94;4: 248-59.
- ^ Hulmes, D. J. S. (2002). „Stavba kolagenních molekul, fibril a suprafibrilárních struktur“. Journal of Structural Biology. 137 (1-2): 2-10. doi:10.1006/jsbi.2002.4450. PMID 12064927.
- ^ Silver, F. H.; Freeman, J. W.; Seehra, G. P. (2003). „Samouspořádání kolagenu a vývoj mechanických vlastností šlach“. Journal of Biomechanics. 36 (10): 1529-1553. doi:10.1016/S0021-9290(03)00135-0. PMID 14499302.
- ^ Ker, R. F. (2002). „Důsledky přizpůsobivé únavové kvality šlach pro jejich konstrukci, opravu a funkci“. Srovnávací biochemie a fyziologie A. 133 (4): 987-1000. doi:10.1016/S1095-6433(02)00171-X. PMID 12485688.
- ^ Cribb, A. M.; Scott, J.E. (1995). In Tendon response to tensile-stress – an ultrastructural investigation of collagen – proteoglycan interactions in stressed tendon,1995; Cambridge Univ Press.pp 423-428.
- ^ Screen H.R., Lee D.A., Bader D.L., Shelton J.C. (2004). „An investigation into the effects of the hierarchical structure of tendon fascicles on micromechanical properties“ (Zkoumání vlivu hierarchické struktury šlachových vazů na mikromechanické vlastnosti). Proc Inst Mech Eng H. 218 (2): 109-119. doi:10.1243/095441104322984004. PMID 15116898. S2CID 46256718. CS1 maint: multiple names: authors list (link)
- ^ Puxkandl, R.; Zizak, I.; Paris, O.; Keckes, J.; Tesch, W.; Bernstorff, S.; Purslow, P.; Fratzl, P. (2002). „Viskoelastické vlastnosti kolagenu: výzkumy pomocí synchrotronového záření a strukturní model“. Philosophical Transactions of the Royal Society B. 357 (1418): 191-197. doi:10.1098/rstb.2001.1033. PMC 1692933. PMID 11911776.
- ^ Gupta H.S., Seto J., Krauss S., Boesecke P.& Screen H.R.C. (2010). Víceúrovňová analýza mechanismů viskoelastické deformace šlachového kolagenu in situ. J. Struct. Biol. 169(2):183-191.
- ^ Thorpe C.T; Udeze C.P; Birch H.L.; Clegg P.D.; Screen H.R.C. (2012). „Specialisation of tendon mechanical properties results from inter-fascicular differences“ (Specializace mechanických vlastností šlach vyplývá z interfascikulárních rozdílů). Journal of the Royal Society Interface. 9 (76): 3108-3117. doi:10.1098/rsif.2012.0362. PMC 3479922. PMID 22764132.
- ^ Thorpe C.T.; Klemt C; Riley G.P.; Birch H.L.; Clegg P.D.; Screen H.R.C. (2013). „Helical sub-structures in energy-storming tendons provide a possible mechanism for efficient energy storage and return“ (Šroubovicové substruktury ve šlachách ukládajících energii představují možný mechanismus pro efektivní ukládání a návrat energie). Acta Biomater. 9 (8): 7948-56. doi:10.1016/j.actbio.2013.05.004. PMID 23669621.
- ^ Gatt R, Vella Wood M, Gatt A, Zarb F, Formosa C, Azzopardi KM, Casha A, Agius TP, Schembri-Wismayer P, Attard L, Chockalingam N, Grima JN (2015). „Negative Poisson’s ratios in tendons: An unexpected mechanical response“. Acta Biomater. 24: 201-208. doi:10.1016/j.actbio.2015.06.018. PMID 26102335.
- ^ Batson EL, Paramour RJ, Smith TJ, Birch HL, Patterson-Kane JC, Goodship AE. (2003). Equine Vet J. |volume=35 |issue=3 |pages=314-8. Jsou materiálové vlastnosti a složení matrice šlach flexorů a extenzorů koní určeny jejich funkcí?
- ^ ScreenH.R.C., Tanner, K.E. (2012). Struktura & Biomechanika biologických kompozitů. In: Sborník abstraktů z konference, která se uskutečnila v roce 2012: Encyclopaedia of Composites 2nd Ed. Nicolais & Borzacchiello.Pub. John Wiley & Sons, Inc. ISBN 978-0-470-12828-2 (strany 2928-39)
- ^ Nakagawa, Y. (1989). „Effect of disuse on the ultra structure of the Achilles tendon in rats“ (Vliv nepoužívání na ultrastrukturu Achillovy šlachy u potkanů). European Journal of Applied Physiology. 59 (3): 239-242. doi:10.1007/bf02386194. PMID 2583169. S2CID 20626078.
- ^ Reeves, N. D. (2005). „Influence of 90-day simulated micro-gravity on human tendon mechanical properties and the effect of restiveness countermeasures“ (Vliv 90denní simulované mikrogravitace na mechanické vlastnosti lidských šlach a účinek protiopatření odpočinku). Journal of Applied Physiology. 98 (6): 2278-2286. doi:10.1152/japplphysiol.01266.2004. hdl:11379/25397. PMID 15705722.
- ^ a b Riley, G. (2004). „Patogeneze tendinopatie. A molecular perspective“ (PDF). Rheumatology. 43 (2): 131-142. doi:10.1093/rheumatology/keg448. PMID 12867575.
- ^ a b c d Sharma, P. M., N. (2006). „Biologie poranění šlach: hojení, modelování a remodelace“. Journal of Musculoskeletal and Neuronal Interactions. 6 (2): 181-190. PMID 16849830. CS1 maint: multiple names: authors list (link)
- ^ a b c d Sharma, P.; Maffulli, N. (2005). „Poranění šlach a tendinopatie: hojení a oprava“. Journal of Bone and Joint Surgery. Americký svazek. 87A (1): 187-202. doi:10.2106/JBJS.D.01850. PMID 15634833. S2CID 1111422.
- ^ a b c d e f Wang, J. H. C. (2006). „Mechanobiologie šlach“. Journal of Biomechanics. 39 (9): 1563-1582. doi:10.1016/j.jbiomech.2005.05.011. PMID 16000201.
- ^ Riley, G. P.; Curry, V.; DeGroot, J.; van El, B.; Verzijl, N.; Hazleman, B. L.; Bank, R. A. (2002). „Matrix metalloproteinase activities and their relationship with collagen remodelling in tendon pathology“ (Aktivity matrixových metaloproteináz a jejich vztah k remodelaci kolagenu v patologii šlach). Matrix Biology. 21 (2): 185-195. doi:10.1016/S0945-053X(01)00196-2. PMID 11852234.
- ^ Moulin, V.; Tam, B. Y. Y.; Castilloux, G.; Auger, F. A.; O’Connor-McCourt, M. D.; Philip, A.; Germain, L. (2001). „Fetální a dospělé lidské kožní fibroblasty vykazují vnitřní rozdíly v kontraktilitě“. Journal of Cellular Physiology. 188 (2): 211-222. doi:10.1002/jcp.1110. PMID 11424088. S2CID 22026692.
- ^ Boyer, M. I. W., J. T.; Lou, J.; Manske, P. R.; Gelberman, R. H.; Cai, S. R. (2001). „Quantitative variation in vascular endothelial growth factor mRNA expression during early flexor tendon healing: an investigation in a canine model“. Journal of Orthopaedic Research. 19 (5): 869-872. doi:10.1016/S0736-0266(01)00017-1. PMID 11562135. S2CID 20903366. CS1 maint: multiple names: authors list (link)
- ^ Astrom, M.; Rausing, A. (1995). „Chronická Achillova tendinopatie – přehled chirurgických a histopatologických nálezů“. Clinical Orthopaedics and Related Research. 316 (316): 151-164. doi:1097/00003086-199507000-00021. PMID 7634699. S2CID 25486134.
- ^ Berge, James C. Vanden; Storer, Robert W. (1995). „Intratendinózní osifikace u ptáků: A review“. Journal of Morphology. 226 (1): 47-77. doi:10.1002/jmor.1052260105. PMID 29865323. S2CID 46926646.
- ^ Organ, Chris L. (2006). „Biomechanika zkostnatělých šlach u ornitopodních dinosaurů“. Paleobiology. 32 (4): 652-665. doi:10.1666/05039.1. S2CID 86568665.