
Cíle učení
- Popsat složky podílející se na svalové kontrakci
- Vysvětlit, jak svaly kontrahují a relaxují
- Popsat model posuvných vláken svalové kontrakce
Sekvence událostí, které vedou ke kontrakci jednotlivých svalových vláken, začíná signálem – neurotransmiterem, ACh z motorického neuronu, který toto vlákno inervuje. Místní membrána vlákna se depolarizuje, protože do ní vstoupí kladně nabité sodíkové ionty (Na+), což spustí akční potenciál, který se rozšíří na zbytek membrány, která se depolarizuje, včetně T-tubulů. To vyvolá uvolnění vápenatých iontů (Ca++) ze zásob v sarkoplazmatickém retikulu (SR). Ca++ pak iniciuje kontrakci, která je udržována ATP (obr. 1). Dokud zůstávají ionty Ca++ v sarkoplazmě, aby se vázaly na troponin, který udržuje vazebná místa aktinu „nezakrytá“, a dokud je k dispozici ATP, který pohání cyklování příčných můstků a tah aktinových vláken myozinem, svalové vlákno se bude nadále zkracovat až na anatomickou mez.

Obrázek 1. Kontrakce svalového vlákna. Mezi aktinem a hlavicemi myozinu vzniká příčný můstek, který spouští kontrakci. Dokud v sarkoplazmě zůstávají ionty Ca++, které se vážou na troponin, a dokud je k dispozici ATP, svalové vlákno se nadále zkracuje.
Svalová kontrakce se obvykle zastaví, když skončí signalizace z motorického neuronu, která repolarizuje sarkolematu a T-tubuly a uzavře napěťově řízené vápníkové kanály v SR. Ionty Ca++ jsou pak pumpovány zpět do SR, což způsobí, že tropomyosin znovu zaštítí (nebo znovu zakryje) vazebná místa na aktinových vláknech. Sval se také může přestat smršťovat, když mu dojde ATP a dojde k jeho únavě (obr. 2).

Obrázek 2. Relaxace svalového vlákna. Ionty Ca++ jsou čerpány zpět do SR, což způsobí, že tropomyosin znovu zaštítí vazebná místa na aktinových vláknech. Sval se také může přestat stahovat, když mu dojde ATP a dojde k únavě.
Molekulární děje při zkracování svalových vláken probíhají uvnitř sarkomer vlákna (viz obrázek 3). Ke kontrakci příčně pruhovaného svalového vlákna dochází tak, že sarkomery, lineárně uspořádané v myofibrilách, se zkracují v důsledku tahu myozinových hlavic za aktinová vlákna.
Oblast, kde se překrývají silná a tenká vlákna, má hustý vzhled, protože mezi vlákny je málo místa. Tato oblast, kde se překrývají tenká a tlustá filamenta, je pro svalovou kontrakci velmi důležitá, protože je to místo, kde začíná pohyb filament. Tenká filamenta, ukotvená na svých koncích pomocí Z-disků, nezasahují zcela do centrální oblasti, která obsahuje pouze silná filamenta, ukotvená u svých základů v místě zvaném M-linie. Myofibrila se skládá z mnoha sarkomer probíhajících po celé její délce; myofibrily a svalové buňky se tedy smršťují tak, jak se smršťují sarkomery.
Model kontrakce posuvnými vlákny
Při signálu z motorického neuronu se kosterní svalové vlákno smršťuje tak, že tenká vlákna jsou tažena a poté se posouvají kolem silných vláken uvnitř sarkomer vlákna. Tento proces je znám jako model svalové kontrakce s posuvnými vlákny (obrázek 3). Ke klouzání může dojít pouze tehdy, když jsou vazebná místa myozinu na aktinových vláknech obnažena sérií kroků, která začíná vstupem Ca++ do sarkoplazmy.

Obrázek 3. Model posuvných vláken při kontrakci svalu. Při kontrakci sarkomery se linie Z přibližují k sobě a pásmo I se zmenšuje. Pás A zůstává stejně široký. Při plné kontrakci se tenká a tlustá vlákna překrývají.
Tropomyozin je protein, který se vine kolem řetězců aktinového vlákna a zakrývá vazebná místa myozinu, aby zabránil vazbě aktinu na myozin. Tropomyosin se váže na troponin a vytváří komplex troponin-tropomyosin. Komplex troponin-tropomyosin brání „hlavičkám“ myosinu vázat se na aktivní místa na aktinových mikrofilamentech. Troponin má také vazebné místo pro ionty Ca++.
Pro zahájení svalové kontrakce musí tropomyosin obnažit vazebné místo myosinu na aktinovém vlákně, aby umožnil vytvoření příčného můstku mezi aktinovými a myosinovými mikrofilamenty. Prvním krokem v procesu kontrakce je vazba Ca++ na troponin, aby se tropomyosin mohl vysunout z vazebných míst na aktinových vláknech. To umožní hlavičkám myozinu navázat se na tato obnažená vazebná místa a vytvořit příčné můstky. Tenká vlákna jsou pak tažena hlavami myozinu, aby se posunula kolem silných vláken směrem do středu sarkomery. Každá hlava však může táhnout jen velmi krátkou vzdálenost, než dosáhne svého limitu a musí se „znovu zakotvit“, aby mohla táhnout znovu, což je krok, který vyžaduje ATP.
ATP a svalová kontrakce
Aby se tenká filamenta mohla během svalové kontrakce dále posouvat kolem silných filament, musí myozinové hlavy táhnout aktin na vazebných místech, odpojit se, znovu zakotvit, připojit se k dalším vazebným místům, táhnout, odpojit se, znovu zakotvit atd. Tento opakovaný pohyb se nazývá cyklus příčných můstků. Tento pohyb myozinových hlavic je podobný pohybu vesel, když jedinec vesluje na lodi: Pádla vesel (myozinové hlavičky) táhnou, jsou zvednuta z vody (odpojena), znovu umístěna (znovu zakotvena) a poté znovu ponořena, aby táhla (obrázek 4). Každý cyklus vyžaduje energii a činnost myozinových hlavic v sarkomerách, které opakovaně tahají za tenká vlákna, také vyžaduje energii, kterou dodává ATP.
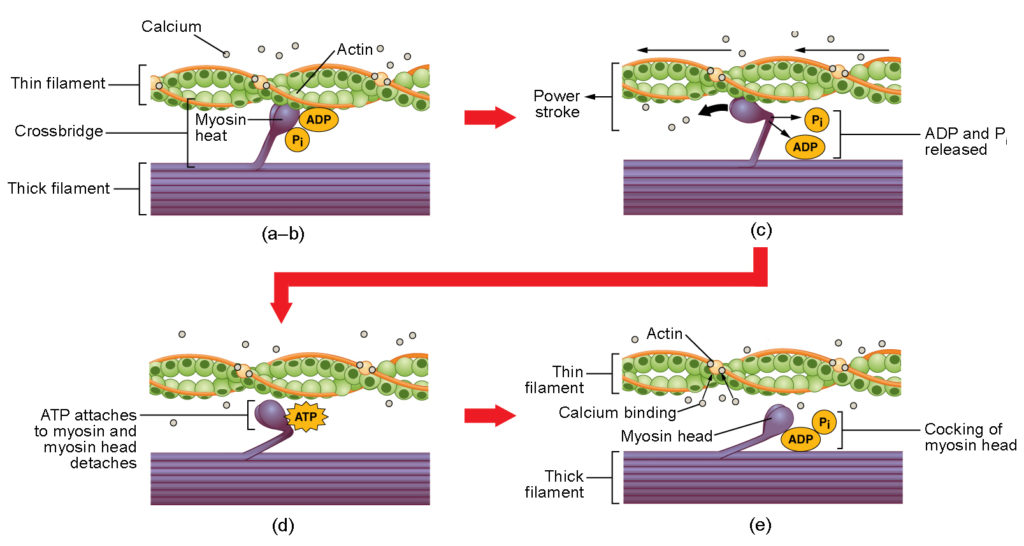
Obrázek 4. Kontrakce kosterního svalu. (a) Aktivní místo na aktinu je odhaleno, když se vápník váže na troponin. (b) Myozinová hlavice je přitahována k aktinu a myozin váže aktin ve svém vazebném místě a vytváří příčný můstek. (c) Během silového tahu se uvolní fosfát vzniklý v předchozím kontrakčním cyklu. To má za následek otočení myozinové hlavy směrem ke středu sarkomery, načež se uvolní navázaný ADP a fosfátová skupina. (d) Na myozinovou hlavici se naváže nová molekula ATP, což způsobí odpojení příčného můstku. (e) Myozinová hlavice hydrolyzuje ATP na ADP a fosfát, čímž se myozin vrátí do kokované polohy.
K tvorbě příčného můstku dochází, když se myozinová hlavice připojí k aktinu, zatímco adenosindifosfát (ADP) a anorganický fosfát (Pi) jsou stále vázány na myozin (obr. 4a,b). Pi se poté uvolní, což způsobí, že myosin vytvoří silnější vazbu k aktinu, načež se hlava myosinu pohybuje směrem k linii M a táhne aktin s sebou. Při tažení aktinu se filamenta posunou přibližně o 10 nm směrem k linii M. Tento pohyb se nazývá silový tah, protože v tomto kroku dochází k pohybu tenkého vlákna (obr. 4c). V nepřítomnosti ATP se myozinová hlavička od aktinu neodpojí.
Jedna část myozinové hlavičky se připojuje k vazebnému místu na aktinu, ale hlavička má další vazebné místo pro ATP. Vazba ATP způsobí, že se hlava myozinu oddělí od aktinu (obrázek 4d). Poté, co k tomu dojde, je ATP přeměněn na ADP a Pi vnitřní ATPázovou aktivitou myosinu. Energie uvolněná při hydrolýze ATP mění úhel myozinové hlavice do vychýlené polohy (obrázek 4e). Hlava myozinu je nyní v poloze vhodné pro další pohyb.
Když je hlava myozinu zakloněná, je myozin ve vysokoenergetické konfiguraci. Tato energie se spotřebovává při pohybu myozinové hlavy v průběhu silového zdvihu a na konci silového zdvihu je myozinová hlava v poloze s nízkou energií. Po skončení silového tahu se ADP uvolní, avšak vytvořený příčný můstek je stále na svém místě a aktin a myozin jsou spojeny dohromady. Dokud je ATP k dispozici, snadno se naváže na myozin, cyklus příčných můstků se může opakovat a svalová kontrakce může pokračovat.
Všimněte si, že každé tlusté vlákno o zhruba 300 molekulách myozinu má více myozinových hlav a během svalové kontrakce se neustále tvoří a rozpadá mnoho příčných můstků. Vynásobte to všemi sarkomérami v jedné myofibrile, všemi myofibrilami v jednom svalovém vlákně a všemi svalovými vlákny v jednom kosterním svalu a pochopíte, proč je k udržení činnosti kosterních svalů potřeba tolik energie (ATP). Ve skutečnosti je to právě ztráta ATP, která má za následek ztuhlost (rigor mortis) pozorovanou brzy poté, co někdo zemře. Protože není možná další produkce ATP, není k dispozici ATP, aby se hlavičky myozinu oddělily od vazebných míst pro aktin, takže příčné můstky zůstávají na místě, což způsobuje ztuhlost kosterních svalů.
Zdroje ATP
ATP dodává energii, aby mohla probíhat svalová kontrakce. Kromě své přímé úlohy v cyklu příčných můstků poskytuje ATP také energii pro aktivní transportní Ca++ pumpy v SR. Ke svalové kontrakci nedochází bez dostatečného množství ATP. Množství ATP uložené ve svalu je velmi nízké, postačuje pouze k pohonu několikavteřinové kontrakce. Protože se ATP odbourává, musí být rychle regenerován a nahrazován, aby byla umožněna trvalá kontrakce. Existují tři mechanismy, kterými lze ATP regenerovat: metabolismus kreatinfosfátu, anaerobní glykolýza, fermentace a aerobní dýchání.
Kreatinfosfát je molekula, která může uchovávat energii ve svých fosfátových vazbách. V klidovém svalu přebytečný ATP předává svou energii kreatinu za vzniku ADP a kreatinfosfátu. Ten funguje jako energetická rezerva, kterou lze využít k rychlé tvorbě dalšího ATP. Když se sval začne stahovat a potřebuje energii, kreatinfosfát převede svůj fosfát zpět na ADP a vytvoří ATP a kreatin. Tato reakce je katalyzována enzymem kreatinkinázou a probíhá velmi rychle; ATP získaný z kreatinfosfátu tak pohání prvních několik sekund svalové kontrakce. Kreatinfosfát však dokáže poskytnout energii v hodnotě přibližně jen 15 sekund, poté je třeba použít jiný zdroj energie (obrázek 5).

Obrázek 5. Svalový metabolismus. V klidovém svalu je uložena část ATP. Jakmile začne kontrakce, spotřebuje se během několika sekund. Další ATP se vytváří z kreatinfosfátu po dobu asi 15 sekund.
Když se ATP produkovaný kreatinfosfátem vyčerpá, svaly se obrátí na glykolýzu jako zdroj ATP. Glykolýza je anaerobní (na kyslíku nezávislý) proces, při kterém se štěpí glukóza (cukr) za vzniku ATP; glykolýza však nemůže vytvářet ATP tak rychle jako kreatinfosfát. Přechod na glykolýzu má tedy za následek pomalejší rychlost dostupnosti ATP pro sval. Cukr používaný při glykolýze může být dodáván z glukózy v krvi nebo metabolizací glykogenu, který je uložen ve svalu. Rozkladem jedné molekuly glukózy vznikají dvě molekuly ATP a dvě molekuly kyseliny pyrohroznové, které mohou být využity při aerobním dýchání nebo při nízkém obsahu kyslíku přeměněny na kyselinu mléčnou (obr. 6).

Obr. 6. Glykolýza a aerobní dýchání. Glykolýza a aerobní dýchání. Každá molekula glukózy produkuje dvě molekuly ATP a dvě molekuly kyseliny pyrohroznové, které mohou být využity při aerobním dýchání nebo přeměněny na kyselinu mléčnou. Pokud není k dispozici kyslík, přeměňuje se kyselina pyrohroznová na kyselinu mléčnou, která může přispívat ke svalové únavě. K tomu dochází při namáhavém cvičení, kdy je potřeba velké množství energie, ale kyslík nemůže být do svalů dostatečně dodáván.
Je-li kyslík k dispozici, kyselina pyrohroznová se využívá při aerobním dýchání. Pokud však kyslík není k dispozici, kyselina pyrohroznová se přeměňuje na kyselinu mléčnou, která může přispívat ke svalové únavě. Tato přeměna umožňuje recyklaci enzymu NAD+ z NADH, který je potřebný pro pokračování glykolýzy. K tomu dochází při namáhavém cvičení, kdy je zapotřebí velké množství energie, ale kyslík nemůže být do svalu dostatečně dodán. Glykolýza sama o sobě nemůže trvat příliš dlouho (přibližně 1 minutu svalové aktivity), ale je užitečná pro usnadnění krátkých dávek vysoce intenzivního výkonu. Je to proto, že glykolýza nevyužívá glukózu příliš efektivně a produkuje čistý zisk dvou ATP na molekulu glukózy a konečný produkt kyseliny mléčné, která může přispívat ke svalové únavě, protože se hromadí.
Aerobní dýchání je rozklad glukózy nebo jiných živin za přítomnosti kyslíku (O2) za vzniku oxidu uhličitého, vody a ATP. Přibližně 95 % ATP potřebného pro klidové nebo středně aktivní svaly je zajištěno aerobním dýcháním, které probíhá v mitochondriích. Vstupními surovinami pro aerobní dýchání jsou glukóza kolující v krevním oběhu, kyselina pyrohroznová a mastné kyseliny. Aerobní dýchání je mnohem účinnější než anaerobní glykolýza a produkuje přibližně 36 ATP na molekulu glukózy oproti čtyřem při glykolýze. Aerobní dýchání však nelze udržet bez stálého přísunu O2 do kosterního svalu a je mnohem pomalejší (obr. 7). Jako kompenzaci ukládají svaly malé množství přebytečného kyslíku do bílkovin zvaných myoglobin, což umožňuje efektivnější svalové kontrakce a menší únavu. Aerobní trénink také zvyšuje účinnost oběhového systému, takže O2 může být do svalů dodáván po delší dobu.

Obrázek 7. Obrázek 7: Svalové dýchání v mitochondriích. Buněčné dýchání. Aerobní dýchání je rozklad glukózy za přítomnosti kyslíku (O2) za vzniku oxidu uhličitého, vody a ATP. Přibližně 95 % ATP potřebného pro klidové nebo středně aktivní svaly je zajištěno aerobním dýcháním, které probíhá v mitochondriích.
Svalová únava nastává, když se sval již nemůže stahovat v reakci na signály z nervového systému. Přesné příčiny svalové únavy nejsou zcela známy, ačkoli určité faktory byly dány do souvislosti se sníženou svalovou kontrakcí, ke které dochází při únavě. Pro normální svalovou kontrakci je zapotřebí ATP, a když se zásoby ATP sníží, může dojít k poklesu svalové funkce. To může být spíše faktorem při krátkém, intenzivním svalovém výkonu než při trvalém úsilí nižší intenzity. Hromadění kyseliny mléčné může snížit intracelulární pH a ovlivnit aktivitu enzymů a bílkovin. Nerovnováha hladin Na+ a K+ v důsledku depolarizace membrán může narušit tok Ca++ ze SR. Dlouhodobé trvalé cvičení může poškodit SR a sarkolemu, což vede k poruše regulace Ca++.
Intenzivní svalová aktivita má za následek kyslíkový dluh, což je množství kyslíku potřebné ke kompenzaci ATP produkovaného bez kyslíku během svalové kontrakce. Kyslík je potřebný k obnovení hladiny ATP a kreatinfosfátu, k přeměně kyseliny mléčné na kyselinu pyrohroznovou a v játrech k přeměně kyseliny mléčné na glukózu nebo glykogen. Kyslík potřebují i další systémy používané během cvičení a všechny tyto kombinované procesy mají za následek zvýšenou dechovou frekvenci, ke které dochází po cvičení. Dokud není kyslíkový dluh vyrovnán, je spotřeba kyslíku zvýšená i po ukončení cvičení.
Relaxace kosterního svalu
Relaxace kosterních svalových vláken a nakonec i kosterního svalu začíná u motorického neuronu, který přestane uvolňovat svůj chemický signál, ACh, do synapse v NMJ. Svalové vlákno se repolarizuje, čímž se uzavřou brány v SR, kde se uvolňoval Ca++. Čerpadla poháněná ATP přesunou Ca++ ze sarkoplazmy zpět do SR. To má za následek „znovuzaštítění“ vazebných míst aktinu na tenkých vláknech. Bez schopnosti vytvářet příčné můstky mezi tenkými a tlustými filamenty ztrácí svalové vlákno napětí a relaxuje.
Svalová síla
Počet vláken kosterního svalu v daném svalu je dán geneticky a nemění se. Svalová síla přímo souvisí s množstvím myofibril a sarkomer v jednotlivých vláknech. Faktory, jako jsou hormony a stres (a umělé anabolické steroidy), působící na sval, mohou zvýšit produkci sarkomer a myofibril uvnitř svalových vláken, což je změna nazývaná hypertrofie, jejímž výsledkem je zvýšení hmotnosti a objemu kosterního svalu. Stejně tak má snížené používání kosterního svalu za následek atrofii, při níž mizí počet sarkomer a myofibril (nikoli však počet svalových vláken). Je běžné, že u končetiny v sádře se po sejmutí sádry projeví atrofie svalů a u některých onemocnění, například u dětské mozkové obrny, se projeví atrofie svalů.
Poruchy svalové soustavy
Duchennova svalová dystrofie (DMD) je progresivní oslabení kosterních svalů. Jedná se o jedno z několika onemocnění souhrnně označovaných jako „svalová dystrofie“. DMD je způsobena nedostatkem bílkoviny dystrofinu, která pomáhá tenkým vláknům myofibril vázat se na sarkolemu. Bez dostatečného množství dystrofinu dochází při svalových kontrakcích k roztržení sarkolemy, což způsobuje příliv Ca++, který vede k poškození buněk a degradaci svalových vláken. Postupem času, jak se poškození svalů hromadí, dochází ke ztrátě svalové hmoty a rozvoji větších funkčních poruch.
DMD je dědičné onemocnění způsobené abnormálním X chromozomem. Postihuje především muže a obvykle je diagnostikována v raném dětství. DMD se obvykle nejprve projevuje jako potíže s rovnováhou a pohybem a poté přechází v neschopnost chodit. Dále postupuje směrem vzhůru po těle od dolních končetin k horní části těla, kde postihuje svaly zodpovědné za dýchání a krevní oběh. Nakonec způsobuje smrt v důsledku respiračního selhání a postižení se obvykle nedožijí více než 20 let.
Protože DMD je způsobena mutací v genu, který kóduje dystrofin, předpokládalo se, že účinnou léčbou by mohlo být vpravení zdravých myoblastů pacientům. Myoblasty jsou embryonální buňky zodpovědné za vývoj svalů a v ideálním případě by nesly zdravé geny, které by mohly produkovat dystrofin potřebný pro normální svalovou kontrakci. Tento přístup byl u lidí většinou neúspěšný. Nedávný přístup spočíval ve snaze zvýšit ve svalu produkci utrofinu, bílkoviny podobné dystrofinu, která by mohla být schopna převzít úlohu dystrofinu a zabránit poškození buněk.
Otázky pro sebekontrolu
Vyzkoušejte si níže uvedený kvíz a ověřte si, zda jste porozuměli tématu Kontrakce a relaxace svalových vláken: