
Kasvot kasvoista kasvoihin on se tapa, jolla useimmiten tunnistamme toisemme ja kommunikoimme keskenämme. Yksittäisten kasvojen tunnistaminen on mahdollista vain siksi, että ihmisen kasvot ovat niin valtavan vaihtelevat. Identtisten kaksosten, jotka perivät kummaltakin vanhemmaltaan samat versiot kustakin geenistä ja joilla on siten identtiset genotyypit, kasvojen äärimmäinen samankaltaisuus osoittaa, että erilaiset kasvonpiirteet, joiden perusteella tunnistamme ihmisiä, ovat periytyviä. Tämä tarkoittaa, että ne määräytyvät enimmäkseen vanhemmilta perittyjen geenimuunnosten erityisten yhdistelmien perusteella. Geneettisellä variantilla tarkoitamme tietyn geenin versiota, joka eroaa saman geenin muista versioista tietyssä DNA:n kohdassa. Se, että erillään kasvaneiden identtisten kaksosten kasvonpiirteet ovat yhtä samankaltaiset kuin yhdessä kasvaneiden, tukee vahvasti näkemystä, jonka mukaan ympäristön vaikutukset kasvonpiirteisiin ovat yleensä hyvin vähäisiä.
Kasvonpiirteet, kuten nenän muoto, taantuva leuka tai ”hapsburgilainen huuli”, periytyvät perheissä usein sukupolvelta toiselle. Tavoitteenamme on ollut tunnistaa erityisiä geneettisiä variantteja, jotka määräävät tietyt kasvonpiirteet. Onnistumisemme tässä tehtävässä, jota ei ole ennen tehty, on riippunut siitä, että olemme käyttäneet monimutkaisia tilastollisia menetelmiä analysoidaksemme vapaaehtoisilta ihmisiltä otettuja kasvokuvia.
Lähtökohtanamme oli suuri kokoelma kolmiulotteisia kasvokuvia, jotka oli otettu huipputeknologian kaupallisella kameralla. Näiden kuvien avulla saimme jonkinlaisen manipuloinnin jälkeen määriteltyä jokaisen kasvon 30 000 pisteeksi kasvojen pinnalla, mikä oli käytännössä kolmiulotteinen kartta kasvoista. Vertaillaksemme eri kasvojen piirteitä asetimme kaikki kasvokuvat päällekkäin toisiinsa nähden. Tämä tehdään siten, että kuvat asetetaan ensin päällekkäin joidenkin kiintopisteiden, kuten nenän kärjen, silmien tai suun kulmien, suhteen. Tämä menettely muistuttaa Francis Galtonin lähes 150 vuotta sitten käyttämää menettelyä, joka oli kasvojen ja kaksostutkimusten uranuurtaja, mutta nykyään meillä on kehittyneitä tietokonetyökaluja ja huipputeknologiaa hyödyntäviä kameroita, jotka parantavat huomattavasti kaikkien kuvien päällekkäisyyttä toisiinsa.
Olemme käyttäneet vapaaehtoisia kolmesta lähteestä: a) 1832 yksilöllistä vapaaehtoista erittäin hyvin luonnehditusta People of the British Isles (PoBI) -tutkimuksestamme, b) 1567 yksilöllistä kaksosta TwinsUK-kohortista, joissa oli suunnilleen yhtä paljon identtisiä ja ei-identtisiä kaksosia, ja c) 33 kuvaa itäaasialaisista, pääasiassa kiinalaisista.
Kaksosten kasvokuvien saaminen mahdollisti analyysin seuraavan tärkeän askeleen, nimittäin kasvonpiirteiden tunnistamisen, joilla on todennäköisesti korkea periytyvyys. Kahdella yksilöllä, jotka ovat identtisiä kaksosia, on samat geneettiset variantit (DNA-sekvenssit). Juuri nämä variantit määräävät heidän kasvonpiirteensä ja johtavat hyvin samankaltaisiin kasvoihin. Kasvonpiirteet määrittyvät kasvojen toisiinsa liittyvien pistemäisten ryhmien avulla, jotka ovat pikemminkin kuin mäet ja laaksot kolmiulotteisella kartalla.
Identtisen kaksosparin toisen jäsenen kasvokuvassa olevan pisteen sijainnin pitäisi olla hyvin samankaltainen kuin toisen kaksosparin kasvokuvassa olevan vastaavan pisteen sijainnin. Se, missä määrin se on erilainen, on mittari niille ei-geneettisille ympäristövaikutuksille, jotka vaikuttavat tämän pisteen sijaintiin kasvoilla. Sitä vastoin kahdella henkilöllä, jotka eivät ole identtisiä kaksosia, voi olla erilaisia geneettisiä variantteja, jotka määrittävät ainakin osan heidän kasvonpiirteistään. Saman pisteen sijainti toisen yksilön, joka ei ole identtinen kaksonen, kasvokuvassa ei siis yleensä ole yhtä lähellä vastaavan pisteen sijaintia toisen kaksosen kuvassa kuin jos he olisivat identtisiä kaksosia. Se, missä määrin pisteet ovat kauempana toisistaan ei-identtisillä kaksosilla kuin identtisillä kaksosilla, on tähän pisteeseen kohdistuvien geneettisten vaikutusten mitta, jota geneetikot kutsuvat periytyvyydeksi. Käyttämällä monimutkaisempia tilastollisia menetelmiä voimme painottaa jokaista kasvojen pistettä sen tällä tavoin mitatulla periytyvyydellä.
Tämän painotuksen vaikutus näkyy kuviossa 1, jossa on piirretty niiden kasvoprofiilin pisteiden frekvenssit, joilla on erilaiset periytyvyydet. Minkä tahansa kohdan periytyvyyden aste vaihtelee arvosta 1, jos mittaustulos on aina täsmälleen sama identtisten kaksosten pareilla, mutta erilainen
ei-identtisten kaksosten pareilla, arvoon 0, jos identtisten kaksosten väliset erot ovat samat kuin ei-identtisten kaksosten väliset erot ja siten käytännössä kaikki
ei-geneettisiä, ensisijaisesti ympäristöstä johtuvia. Punaiset sarakkeet kuvaavat painotettuja arvoja, siniset alkuperäisiä arvoja ja violetit päällekkäisyyksiä. Punainen profiili on keskimäärin selvästi korkeampi ja paljon kapeampi kuin sininen, mikä osoittaa painotuksen suotuisan vaikutuksen.
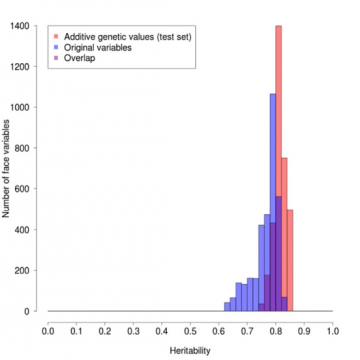
Kuvio 1: Profiilin periytyvyyksien vertailu alkuperäisten ja periytyvyyspainotettujen arvojen välillä.
Seuraavana haasteena on määritellä geneettisessä analyysissä käytettävät kasvonpiirteet toisiinsa assosioituneiden pisteiden ryhmien perusteella. Tätä varten käytämme periytyvyyspainotettuja pisteitä olettaen, että näin saadaan piirre, joka on kaiken kaikkiaan todennäköisesti periytyvämpi kuin painottamattomien pisteiden avulla saatu piirre. Painotettuja pisteitä käytettiin tilastotieteilijät kutsuvat PCA:ksi, joka tarkoittaa pääkomponenttianalyysia. Tämä on tapa poimia aineistosta ne piirteet, jotka ovat eniten muuttuvia. Jokainen PCA, ja niitä voi olla jopa 50 tai useampia, jos kyseessä on monimutkainen data, kuten kasvokuvat, on itsessään kasvonpiirteen mitta, joka on analoginen silmien välisen etäisyyden kanssa, mutta joka yhdistää tehokkaasti eri mittaukset, jotka perustuvat pistejoukkoon, yhdeksi ainoaksi arvoksi.
Tähän mennessä kasvonpiirteiden määrittelyssä emme ole käyttäneet erityistä geneettistä tietoa. Meillä on DNA:han perustuvaa geneettistä tietoa noin 500 000 variantista jokaisesta noin 1500 henkilöstä PoBI-vapaaehtoisistamme, joista meillä on kuvia, ja vastaavasta määrästä TwinsUK-vapaaehtoisia, joista meillä on kuvia. Seuraava vaihe oli siis etsiä erityisiä geneettisiä variantteja, jotka liittyivät merkittävästi PCA:han perustuviin kasvonpiirteisiimme.
Lähestymistapamme geneettiseen analyysiin perustuu ajatukseen, että kasvonpiirteiden eroja olisi analysoitava erillisinä, yksilöllisesti tunnistettavina ominaisuuksina eikä määrällisenä mittana, kuten henkilön pituus. Emme voi tunnistaa ihmistä pelkästään hänen pituutensa tai minkään yksittäisen kvantitatiivisen kasvonpiirteen, kuten silmien välisen etäisyyden tai kasvojen korkeuden ja leveyden suhteen perusteella. Tätä käsitellään keskittymällä niihin henkilöihin, jotka ovat kunkin PCA:n antaman arvon äärimmäisessä ylä- tai alapuolella (10 %), ja kysymällä, onko heillä enemmän yhteistä yhtä tai useampaa 500 000 geneettisestä markkerista kuin niillä henkilöillä, jotka eivät ole näissä ääripäissä. Käytimme People of the British Isles -vapaaehtoisia valitsemaan 500 000 testatusta geenivariantista ehdokkaita jatkoanalyyseihin ääripäiden ja ei-ääripäiden välisen eron merkitsevyyden ja tämän eron suuruuden perusteella. Tämän jälkeen kysyimme, oliko jokin näistä ehdokkaiden vaikutuksista toistunut noin 1500 TwinsUK-vapaaehtoisella. Tällä tavoin olemme tunnistaneet kolme spesifistä ja toistettua geneettistä varianttia, joilla on suhteellisen suuria vaikutuksia, kaksi kasvoprofiilin piirteisiin ja yksi silmien ympärillä olevaan alueeseen. Kullakin näistä kolmesta variantista on
kumppani, jolla on eri DNA-sekvenssi samassa kriittisessä kohdassa, ja kussakin tapauksessa positiivisesti assosioituneen variantin PoBI-populaatiotaajuus on noin 10 %, kun taas kumppanilla on korkeampi, noin 90 %:n taajuus. Kutsumme assosioitunutta varianttia a:ksi ja sen kumppania A:ksi, ja koska geenit ovat pareittain, näitä variantteja on kolme yhdistelmää, aa, Aa ja AA.
Ensimmäinen näistä varianteista, joka löydettiin PCDH15-nimisestä geenistä, lisäsi todennäköisyyttä saada kuvassa 2C esitetyt naisen piirteet yli 7-kertaiseksi niillä brittiläisillä naisilla, jotka kantoivat molempia kopioita varianttia (aa), verrattuna niihin naisiin, joilla oli varianttia vain yksi (Aa) tai ei yhtään kopiota (AA). Tämä variantti liittyy myös piirteisiin, jotka eroavat brittiläisten ja itäaasialaisten vapaaehtoisten naisten välillä. Huomaa nenän ja ylähuulen ylöspäin kääntynyt pää ja vetäytynyt leuka kuvassa 2A, joka on kiinalaisten kasvojen keskiarvo, ja kuvassa 2B, joka on PoBI-henkilöiden kiinalaisempi ryhmä, ja aseta tämä vastakkain kuvan 2C kanssa. PCDH15-geenin tuotetta löytyy kehittyvien hiirten nenän hajusoluista ja rustosta, mikä on sopusoinnussa
ihmisillä löytämämme muunnoksen mahdollisen nenään kohdistuvan vaikutuksen kanssa.
Toinen muunnos, MBTPS1-nimisessä geenissä, liittyy kuvassa 3 esitettyyn kasvojen eroon. Tämä ero havaittiin naisilla, ja erottuva osajoukko kasvoja kantoi molempia kopioita variantista (aa). Tässä tapauksessa ylempään ääripään fenotyyppiin (kuva 3A) liittyvä geenivariantti esiintyy (oletettavasti aa:na) afrikkalaisessa vihreässä apinassa, makakissa ja oliivipaviaanissa, kun taas sen kumppani, yhteinen variantti, esiintyy (oletettavasti AA:na) orangutanissa, gorillassa, simpanssissa ja marsimetsissä, mikä viittaa siihen, että tämä varianttiero saattaa liittyä näiden kädellisryhmien välisiin kasvojen eroihin.
Kolmas variantti, geenissä nimeltä TMEM163, liittyy molemmilla sukupuolilla silmien eroihin, kuten kuvassa 4 on esitetty. Tämän geenin viallisella versiolla on mahdollinen rooli sairaudessa nimeltä mukolipidoosi tyyppi IV, sairaus, johon toisinaan liittyy kasvojen poikkeavuuksia, erityisesti silmäluomien ympärillä. Tutkimuksissamme juuri niiden henkilöiden osajoukko, jotka kantavat muunnoksen (aa) molempia kopioita, liittyy kuvassa 4A esitettyyn yläääriin. Huomaa, että silmien leveys ja silmien korkeus (kulmakarvojen alareunasta silmäluomen yläreunaan) ovat molemmat suuremmat ylemmässä ääripäässä kuin alemmassa ääripäässä.
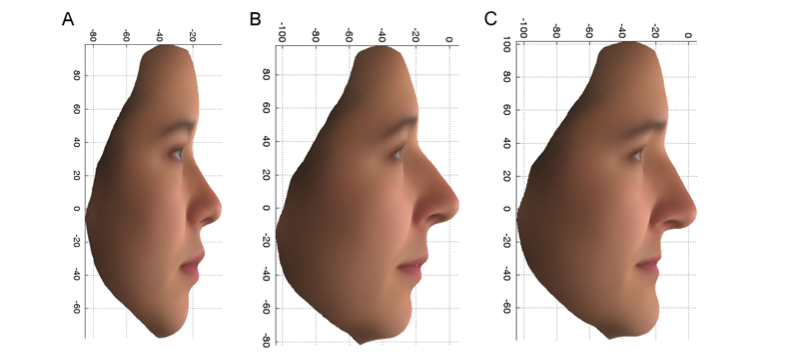
Kuva 2: PC2-profiili: Keskimääräiset kasvot alkuperäisiä muuttujia käyttäen 14 itäaasialaiselle naiselle (A) ja PoBI-naisten ylemmät 10 % (enemmän itäaasialaisia) (B) ja alemmat 10 % (enemmän eurooppalaisia) (C) ääripäät.
Jokainen kolmesta geneettisestä variantista, jotka olemme pystyneet yhdistämään tiettyyn kasvonpiirteeseen, lisää todennäköisyyttä saada kyseinen piirre
yli 7-kertaisesti niillä brittiläisillä vapaaehtoisilla, jotka kantavat variantin molempia kopioita (aa), verrattuna niihin, joilla on vain toinen (Aa) tai ei yhtään (AA) kopiota variantista.

Kuva 3: PC7-profiili: Naisten kasvojen keskimääräiset profiilit alkuperäisiä muuttujia käyttäen varianttiin liittyvien ylempien 10 %:n (A) ja alempien 10 %:n (C) ääripäiden sekä kokonaiskeskiarvon (B) osalta.

Kuva 4: PC1-silmät: Keskimääräiset silmäfenotyypit alkuperäisiä muuttujia käyttäen ylemmän 10 %:n (A), alemman 10 %:n (C) ääripäiden ja kokonaiskeskiarvon (B) osalta.
Onnistumisemme näiden geneettisten varianttien löytämisessä riippuu suurelta osin kyvystämme tunnistaa kasvonpiirteet, joilla on korkea periytyvyys kaksosdatan perusteella, ja ääripäiden valinnasta geneettisten varianttien yhteyksien tutkimiseksi. Näyttää todennäköiseltä, että tulevaisuudessa löydetään paljon enemmän spesifisiä ja suhteellisen suuria geneettisten varianttien vaikutuksia ihmisen kasvonpiirteisiin käyttämällä kuvaamiemme kaltaisia lähestymistapoja. Tämä tasoittaa tietä kohti niiden molekulaaristen mekanismien selvittämistä, joiden avulla geneettiset variantit määrittävät ihmisen kasvojen ulkonäön poikkeuksellista vaihtelua.