
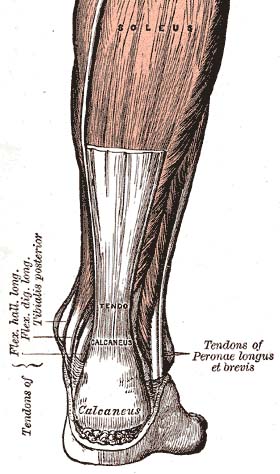

tendo
D013710
H3.03.00.0.00020
Terminologia anatomica
Un tendine è una banda resistente di tessuto connettivo fibroso che collega il muscolo all’osso ed è capace di sopportare la tensione.
I tendini sono simili ai legamenti; entrambi sono fatti di collagene. I legamenti collegano un osso all’altro, mentre i tendini collegano il muscolo all’osso.
Struttura
I tendini sono costituiti da tessuto connettivo denso e regolare. Il principale componente cellulare dei tendini sono fibroblasti specializzati chiamati tenociti. I tenociti sintetizzano la matrice extracellulare dei tendini, abbondante di fibre di collagene densamente imballate. Le fibre di collagene sono parallele tra loro e organizzate in fascicoli. I singoli fascicoli sono legati dall’endotendineo, che è un delicato tessuto connettivo sciolto contenente sottili fibrille di collagene e fibre elastiche. I gruppi di fascicoli sono legati dall’epitenone, che è una guaina di denso tessuto connettivo irregolare. L’intero tendine è racchiuso da una fascia. Lo spazio tra la fascia e il tessuto tendineo è riempito dal paratenon, un tessuto grasso areolare. I normali tendini sani sono ancorati all’osso dalle fibre di Sharpey.
Matrice extracellulare
La massa secca dei tendini normali, che costituisce il 30-45% della loro massa totale, è composta da:
- 60-85% collagene
- 60-80% collagene I
- 0-10% collagene III
- 2% collagene IV
- piccole quantità di collageni V, VI, e altri
- 15-40% componenti della matrice extracellulare non collagene, compresi:
- 3% proteina della matrice oligomerica della cartilagine,
- 1-2% elastina,
- 1-5% proteoglicani,
- 0.2% di componenti inorganici come rame, manganese e calcio.
Mentre il collagene I costituisce la maggior parte del collagene nel tendine, sono presenti molti collageni minori che svolgono ruoli vitali nel corretto sviluppo e funzione del tendine. Questi includono il collagene di tipo II nelle zone cartilaginee, il collagene di tipo III nelle fibre di reticolina delle pareti vascolari, il collagene di tipo IX, il collagene di tipo IV nelle membrane basali dei capillari, il collagene di tipo V nelle pareti vascolari e il collagene di tipo X nel fibrocartilagine mineralizzato vicino all’interfaccia con l’osso.
Ultrastruttura e sintesi del collagene
Le fibre di collagene si fondono in macroaggregati. Dopo la secrezione dalla cellula, scissa dalle proteasi N e C del procollagene, le molecole di tropocollagene si assemblano spontaneamente in fibrille insolubili. Una molecola di collagene è lunga circa 300 nm e larga 1-2 nm, e il diametro delle fibrille che si formano può variare da 50-500 nm. Nei tendini, le fibrille si assemblano ulteriormente per formare fascicoli, che sono lunghi circa 10 mm con un diametro di 50-300 μm, e infine in una fibra tendinea con un diametro di 100-500 μm.
Il collagene dei tendini è tenuto insieme da componenti proteoglicani (un composto costituito da una proteina legata a gruppi glicosaminoglicani, presente soprattutto nel tessuto connettivo) tra cui la decorina e, nelle regioni compresse del tendine, l’aggrecano, che sono in grado di legarsi alle fibrille di collagene in punti specifici. I proteoglicani sono intrecciati con le fibrille di collagene – le loro catene laterali di glicosaminoglicani (GAG) hanno interazioni multiple con la superficie delle fibrille – dimostrando che i proteoglicani sono importanti strutturalmente nell’interconnessione delle fibrille. I principali componenti GAG del tendine sono il solfato di dermatano e il solfato di condroitina, che si associano al collagene e sono coinvolti nel processo di assemblaggio delle fibrille durante lo sviluppo del tendine. Si pensa che il solfato di dermatano sia responsabile della formazione di associazioni tra le fibrille, mentre il solfato di condroitina è più coinvolto nell’occupare il volume tra le fibrille per tenerle separate e aiutare a resistere alla deformazione. Le catene laterali di dermatan solfato della decorina si aggregano in soluzione e questo comportamento può aiutare l’assemblaggio delle fibrille di collagene. Quando le molecole di decorina sono legate a una fibrilla di collagene, le loro catene di dermatan solfato possono estendersi e associarsi con altre catene di dermatan solfato sulla decorina che è legata a fibrille separate, creando così ponti interfibrillari e causando alla fine l’allineamento parallelo delle fibrille.
Tenociti
I tenociti producono le molecole di collagene, che si aggregano da un capo all’altro e da un lato all’altro per produrre fibrille collagene. I fasci di fibrille sono organizzati per formare fibre con i tenociti allungati strettamente imballati tra di loro. C’è una rete tridimensionale di processi cellulari associati al collagene nel tendine. Le cellule comunicano tra loro attraverso le gap junctions, e questa segnalazione dà loro la capacità di rilevare e rispondere al carico meccanico.
I vasi sanguigni possono essere visualizzati all’interno dell’endotendine che corre parallelo alle fibre di collagene, con occasionali anastomosi trasversali ramificate.
Si pensa che la massa interna del tendine non contenga fibre nervose, ma l’epitenone e il paratenone contengono terminazioni nervose, mentre gli organi tendinei del Golgi sono presenti alla giunzione tra tendine e muscolo.
La lunghezza del tendine varia in tutti i principali gruppi e da persona a persona. La lunghezza del tendine è, in pratica, il fattore decisivo per quanto riguarda le dimensioni reali e potenziali del muscolo. Per esempio, a parità di altri fattori biologici rilevanti, un uomo con un tendine più corto e un muscolo bicipite più lungo avrà un potenziale di massa muscolare maggiore di un uomo con un tendine più lungo e un muscolo più corto. I bodybuilder di successo avranno generalmente tendini più corti. Al contrario, negli sport che richiedono agli atleti di eccellere in azioni come la corsa o il salto, è vantaggioso avere un tendine d’Achille più lungo della media e un muscolo del polpaccio più corto.
La lunghezza del tendine è determinata dalla predisposizione genetica, e non è stato dimostrato che aumenta o diminuisce in risposta all’ambiente, a differenza dei muscoli, che possono essere accorciati da traumi, squilibri d’uso e una mancanza di recupero e stretching. RAT
Funzioni

Tradizionalmente, i tendini sono stati considerati come un meccanismo attraverso il quale i muscoli si collegano alle ossa così come i muscoli stessi, funzionando per trasmettere forze. Questa connessione permette ai tendini di modulare passivamente le forze durante la locomozione, fornendo ulteriore stabilità senza lavoro attivo. Tuttavia, negli ultimi due decenni, molte ricerche si sono concentrate sulle proprietà elastiche di alcuni tendini e sulla loro capacità di funzionare come molle. Non tutti i tendini devono svolgere lo stesso ruolo funzionale, con alcuni che posizionano prevalentemente gli arti, come le dita quando si scrive (tendini posizionali) e altri che fungono da molle per rendere la locomozione più efficiente (tendini accumulatori di energia). I tendini accumulatori di energia possono immagazzinare e recuperare energia ad alta efficienza. Per esempio, durante un passo umano, il tendine di Achille si allunga mentre l’articolazione della caviglia si dorsiflette. Durante l’ultima parte del passo, quando il piede si flette plantarmente (puntando le dita verso il basso), l’energia elastica immagazzinata viene rilasciata. Inoltre, poiché il tendine si allunga, il muscolo è in grado di funzionare con meno o addirittura nessuna variazione di lunghezza, permettendo al muscolo di generare più forza.
Le proprietà meccaniche del tendine dipendono dal diametro e dall’orientamento delle fibre di collagene. Le fibrille di collagene sono parallele l’una all’altra e strettamente imballate, ma mostrano un aspetto ondulatorio dovuto a ondulazioni planari, o pieghe, su una scala di diversi micrometri. Nei tendini, le fibre di collagene hanno una certa flessibilità dovuta all’assenza di residui di idrossiprolina e prolina in punti specifici della sequenza aminoacidica, il che permette la formazione di altre conformazioni come curve o anelli interni nella tripla elica e provoca lo sviluppo delle pieghe. Le pieghe nelle fibrille di collagene permettono ai tendini di avere una certa flessibilità e una bassa rigidità alla compressione. Inoltre, poiché il tendine è una struttura a più fili composta da molte fibrille e fascicoli parzialmente indipendenti, non si comporta come una singola barra, e questa proprietà contribuisce anche alla sua flessibilità.
Anche i componenti proteoglicani dei tendini sono importanti per le proprietà meccaniche. Mentre le fibrille di collagene permettono ai tendini di resistere alla trazione, i proteoglicani permettono loro di resistere alla compressione. Queste molecole sono molto idrofile, il che significa che possono assorbire una grande quantità di acqua e quindi hanno un alto rapporto di rigonfiamento. Poiché sono legati in modo non covalente alle fibrille, possono associarsi e dissociarsi reversibilmente in modo che i ponti tra le fibrille possano essere rotti e riformati. Questo processo può essere coinvolto nel permettere alla fibrilla di allungarsi e diminuire di diametro sotto tensione. Tuttavia, i proteoglicani possono anche avere un ruolo nelle proprietà tensili del tendine. La struttura del tendine è effettivamente un materiale composito di fibre, costruito come una serie di livelli gerarchici. Ad ogni livello della gerarchia, le unità di collagene sono legate insieme da legami incrociati di collagene o dai proteoglicani, per creare una struttura altamente resistente al carico di trazione. È stato dimostrato che l’allungamento e la deformazione delle sole fibrille di collagene sono molto inferiori all’allungamento e alla deformazione totali dell’intero tendine sotto la stessa quantità di stress, dimostrando che anche la matrice ricca di proteoglicani deve subire una deformazione, e l’irrigidimento della matrice avviene ad alti tassi di deformazione. Questa deformazione della matrice non collagene si verifica a tutti i livelli della gerarchia del tendine, e modulando l’organizzazione e la struttura di questa matrice, si possono ottenere le diverse proprietà meccaniche richieste dai diversi tendini. È stato dimostrato che i tendini ad accumulo di energia utilizzano quantità significative di scorrimento tra i fascicoli per consentire le caratteristiche di alta deformazione che richiedono, mentre i tendini posizionali si basano più pesantemente sullo scorrimento tra le fibre di collagene e le fibrille. Tuttavia, dati recenti suggeriscono che i tendini ad accumulo di energia possono anche contenere fascicoli che sono contorti, o elicoidali, in natura – una disposizione che sarebbe altamente vantaggiosa per fornire il comportamento elastico richiesto in questi tendini.
Meccanica
I tendini sono strutture viscoelastiche, il che significa che mostrano un comportamento sia elastico che viscoso. Quando vengono allungati, i tendini mostrano un tipico comportamento da “tessuto molle”. La curva forza-estensione, o stress-strain, inizia con una regione di rigidità molto bassa, poiché la struttura di crimpatura si raddrizza e le fibre di collagene si allineano suggerendo un rapporto di Poisson negativo nelle fibre del tendine. Più recentemente, i test effettuati in vivo (attraverso la risonanza magnetica) ed ex vivo (attraverso test meccanici di vari tessuti tendinei cadaverici) hanno dimostrato che i tendini sani sono altamente anisotropi ed esibiscono un rapporto di Poisson negativo (auxetico) in alcuni piani quando vengono allungati fino al 2% della loro lunghezza, cioè entro il loro normale range di movimento. Dopo questa regione di “punta”, la struttura diventa significativamente più rigida, e ha una curva sforzo-deformazione lineare fino a quando non inizia a cedere. Le proprietà meccaniche dei tendini variano ampiamente, in quanto sono abbinate ai requisiti funzionali del tendine. I tendini che immagazzinano energia tendono ad essere più elastici, o meno rigidi, in modo da poter immagazzinare più facilmente energia, mentre i tendini posizionali più rigidi tendono ad essere un po’ più viscoelastici, e meno elastici, in modo da poter fornire un controllo più fine del movimento. Un tipico tendine immagazzinatore di energia si romperà a circa il 12-15% di sforzo, e una sollecitazione nella regione di 100-150 MPa, anche se alcuni tendini sono notevolmente più estensibili di questo, per esempio il flessore digitale superficiale del cavallo, che si allunga in eccesso del 20% quando si galoppa. I tendini posizionali possono cedere a sforzi fino al 6-8%, ma possono avere moduli nella regione di 700-1000 MPa.
Diversi studi hanno dimostrato che i tendini rispondono ai cambiamenti di carico meccanico con processi di crescita e rimodellamento, proprio come le ossa. In particolare, uno studio ha dimostrato che il disuso del tendine di Achille nei ratti ha portato a una diminuzione dello spessore medio dei fasci di fibre di collagene che compongono il tendine. Negli esseri umani, un esperimento in cui le persone sono state sottoposte a un ambiente di microgravità simulata ha scoperto che la rigidità del tendine è diminuita significativamente, anche quando ai soggetti è stato richiesto di eseguire esercizi di riposo. Questi effetti hanno implicazioni in aree che vanno dal trattamento di pazienti costretti a letto alla progettazione di esercizi più efficaci per gli astronauti.
Guarigione
I tendini del piede sono molto complessi e intricati. Pertanto, il processo di guarigione di un tendine rotto è lungo e doloroso. La maggior parte delle persone che non ricevono assistenza medica entro le prime 48 ore dalla lesione, soffrono di un forte gonfiore, dolore e una sensazione di bruciore nel punto in cui si è verificata la lesione.
Si credeva che i tendini non potessero subire il turnover della matrice e che i tenociti non fossero in grado di riparare. Tuttavia, è stato dimostrato che, nel corso della vita di una persona, i tenociti del tendine sintetizzano attivamente i componenti della matrice e gli enzimi come le metalloproteinasi di matrice (MMP) possono degradare la matrice. I tendini sono in grado di guarire e recuperare dalle lesioni in un processo che è controllato dai tenociti e dalla loro matrice extracellulare circostante.
Le tre fasi principali della guarigione dei tendini sono l’infiammazione, la riparazione o proliferazione, e il rimodellamento, che può essere ulteriormente suddiviso in consolidamento e maturazione. Queste fasi possono sovrapporsi l’una all’altra. Nella prima fase, le cellule infiammatorie come i neutrofili vengono reclutate nel sito della lesione, insieme agli eritrociti. I monociti e i macrofagi vengono reclutati entro le prime 24 ore e si verifica la fagocitosi dei materiali necrotici nel sito della lesione. Dopo il rilascio di fattori vasoattivi e chemiotattici, inizia l’angiogenesi e la proliferazione dei tenociti. I tenociti si spostano poi nel sito e iniziano a sintetizzare il collagene III. Dopo alcuni giorni, inizia la fase di riparazione o proliferazione. In questa fase, i tenociti sono coinvolti nella sintesi di grandi quantità di collagene e proteoglicani nel sito della lesione, e i livelli di GAG e acqua sono alti. Dopo circa sei settimane, inizia la fase di rimodellamento. La prima parte di questa fase è il consolidamento, che dura da circa sei a dieci settimane dopo la lesione. Durante questo periodo, la sintesi del collagene e dei GAG è diminuita, e anche la cellularità è diminuita mentre il tessuto diventa più fibroso come risultato di una maggiore produzione di collagene I e le fibrille si allineano nella direzione dello stress meccanico. La fase finale di maturazione si verifica dopo dieci settimane, e durante questo periodo c’è un aumento della reticolazione delle fibrille di collagene, che fa sì che il tessuto diventi più rigido. Gradualmente, nel corso di circa un anno, il tessuto si trasformerà da fibroso a cicatriziale.
Le metalloproteinasi della matrice (MMPs) hanno un ruolo molto importante nella degradazione e nel rimodellamento della ECM durante il processo di guarigione dopo una lesione tendinea. Alcune MMP, tra cui MMP-1, MMP-2, MMP-8, MMP-13 e MMP-14 hanno attività collagenasica, il che significa che, a differenza di molti altri enzimi, sono in grado di degradare le fibrille di collagene I. La degradazione delle fibrille di collagene da parte della MMP-1 insieme alla presenza di collagene denaturato sono fattori che si ritiene causino l’indebolimento dell’ECM del tendine e un aumento del potenziale di un’altra rottura. In risposta a ripetuti carichi meccanici o lesioni, le citochine possono essere rilasciate dai tenociti e possono indurre il rilascio di MMPs, causando la degradazione dell’ECM e portando a lesioni ricorrenti e tendinopatie croniche.
Una varietà di altre molecole sono coinvolte nella riparazione e rigenerazione del tendine. Ci sono cinque fattori di crescita che hanno dimostrato di essere significativamente upregolati e attivi durante la guarigione del tendine: fattore di crescita insulino-simile 1 (IGF-I), fattore di crescita derivato dalle piastrine (PDGF), fattore di crescita endoteliale vascolare (VEGF), fattore di crescita dei fibroblasti di base (bFGF), e fattore di crescita trasformante beta (TGF-β). Questi fattori di crescita hanno tutti ruoli diversi durante il processo di guarigione. L’IGF-1 aumenta la produzione di collagene e proteoglicani durante la prima fase dell’infiammazione, e anche il PDGF è presente durante le prime fasi dopo la lesione e promuove la sintesi di altri fattori di crescita insieme alla sintesi del DNA e la proliferazione delle cellule del tendine. Le tre isoforme di TGF-β (TGF-β1, TGF-β2, TGF-β3) sono note per avere un ruolo nella guarigione delle ferite e nella formazione delle cicatrici. Il VEGF è noto per promuovere l’angiogenesi e per indurre la proliferazione e la migrazione delle cellule endoteliali, ed è stato dimostrato che l’mRNA del VEGF è espresso nel sito delle lesioni tendinee insieme all’mRNA del collagene I. Le proteine morfogenetiche ossee (BMP) sono un sottogruppo della superfamiglia TGF-β che può indurre la formazione di osso e cartilagine così come la differenziazione dei tessuti, e BMP-12 in particolare ha dimostrato di influenzare la formazione e la differenziazione del tessuto tendineo e di promuovere la fibrogenesi.
Effetti dell’attività sulla guarigione
In modelli animali, sono stati condotti studi approfonditi per indagare gli effetti dello sforzo meccanico sotto forma di livello di attività sulle lesioni e sulla guarigione del tendine. Mentre lo stretching può interrompere la guarigione durante la fase infiammatoria iniziale, è stato dimostrato che il movimento controllato dei tendini dopo circa una settimana dopo una lesione acuta può aiutare a promuovere la sintesi di collagene da parte dei tenociti, portando ad un aumento della resistenza alla trazione e del diametro dei tendini guariti e meno aderenze rispetto ai tendini che sono immobilizzati. Nelle lesioni croniche dei tendini, il carico meccanico ha anche dimostrato di stimolare la proliferazione dei fibroblasti e la sintesi del collagene insieme al riallineamento del collagene, tutti fattori che promuovono la riparazione e il rimodellamento. Per sostenere ulteriormente la teoria che il movimento e l’attività aiutano la guarigione dei tendini, è stato dimostrato che l’immobilizzazione dei tendini dopo una lesione ha spesso un effetto negativo sulla guarigione. Nei conigli, i fascicoli di collagene che sono immobilizzati hanno mostrato una minore resistenza alla trazione, e l’immobilizzazione si traduce anche in una minore quantità di acqua, proteoglicani e legami incrociati di collagene nei tendini.
Diversi meccanismi di meccanotrasduzione sono stati proposti come ragioni per la risposta dei tenociti alla forza meccanica che permette loro di alterare la loro espressione genica, la sintesi proteica e il fenotipo cellulare, e infine causare cambiamenti nella struttura del tendine. Un fattore importante è la deformazione meccanica della matrice extracellulare, che può influenzare il citoscheletro di actina e quindi influenzare la forma, la motilità e la funzione delle cellule. Le forze meccaniche possono essere trasmesse da siti di adesione focale, integrine e giunzioni cellula-cellula. I cambiamenti nel citoscheletro di actina possono attivare le integrine, che mediano la segnalazione “outside-in” e “inside-out” tra la cellula e la matrice. Anche le proteine G, che inducono cascate di segnalazione intracellulare, possono essere importanti, e i canali ionici sono attivati dallo stiramento per permettere a ioni come calcio, sodio o potassio di entrare nella cellula.
Società e cultura
Il tendine era ampiamente usato durante le epoche pre-industriali come fibra dura e durevole. Alcuni usi specifici includono l’uso della carbonchio come filo per cucire, per attaccare le piume alle frecce (vedi fletch), per legare le lame degli utensili alle aste, ecc. È anche raccomandato nelle guide di sopravvivenza come materiale da cui si possono fare corde forti per oggetti come trappole o strutture abitative. Il tendine deve essere trattato in modi specifici per funzionare utilmente per questi scopi. Gli Inuit e altre popolazioni circumpolari utilizzavano il tendine come unico cordame per tutti gli scopi domestici a causa della mancanza di altre fonti di fibre adatte nei loro habitat ecologici. Le proprietà elastiche di particolari tendini erano anche usate in archi compositi ricurvi favoriti dai nomadi delle steppe dell’Eurasia e dai nativi americani. Anche la prima artiglieria da lancio di pietre utilizzava le proprietà elastiche del tendine.
Il tendine è un eccellente materiale per cordami per tre motivi: È estremamente forte, contiene colle naturali, e si restringe quando si asciuga, eliminando la necessità di nodi.
Usi culinari
Il tendine (in particolare quello di manzo) è usato come cibo in alcune cucine asiatiche (spesso servito nei ristoranti yum cha o dim sum). Un piatto popolare è il suan bao niu jin, in cui il tendine è marinato in aglio. A volte si trova anche nel piatto vietnamita di noodle phở.
Significato clinico
Lesioni
I tendini sono soggetti a molti tipi di lesioni. Ci sono varie forme di tendinopatie o lesioni tendinee dovute a un uso eccessivo. Questi tipi di lesioni generalmente provocano un’infiammazione e una degenerazione o un indebolimento dei tendini, che alla fine può portare alla rottura del tendine. Le tendinopatie possono essere causate da una serie di fattori relativi alla matrice extracellulare del tendine (ECM), e la loro classificazione è stata difficile perché i loro sintomi e la loro istopatologia sono spesso simili.
La prima categoria di tendinopatia è la paratenonite, che si riferisce all’infiammazione del paratenone, o foglio paratendineo situato tra il tendine e la sua guaina. La tendinosi si riferisce alla lesione non infiammatoria del tendine a livello cellulare. La degradazione è causata da danni al collagene, alle cellule e ai componenti vascolari del tendine, ed è nota per portare alla rottura. Le osservazioni dei tendini che hanno subito una rottura spontanea hanno mostrato la presenza di fibrille di collagene che non sono nel corretto orientamento parallelo o non sono uniformi in lunghezza o diametro, insieme a tenociti arrotondati, altre anomalie cellulari e la crescita di vasi sanguigni. Altre forme di tendinosi che non hanno portato alla rottura hanno anche mostrato la degenerazione, il disorientamento e l’assottigliamento delle fibrille di collagene, insieme ad un aumento della quantità di glicosaminoglicani tra le fibrille. La terza è la paratenonite con tendinosi, in cui sono presenti combinazioni di infiammazione del paratenone e degenerazione del tendine. L’ultima è la tendinite, che si riferisce alla degenerazione con infiammazione del tendine così come la rottura vascolare.
Le tendinopatie possono essere causate da diversi fattori intrinseci tra cui l’età, il peso corporeo e l’alimentazione. I fattori estrinseci sono spesso legati allo sport e includono forze o carichi eccessivi, tecniche di allenamento inadeguate e condizioni ambientali.
Altri animali

In alcuni organismi, tra cui spiccano gli uccelli e i dinosauri ornitischi, porzioni del tendine possono ossificarsi. In questo processo, gli osteociti si infiltrano nel tendine e depongono l’osso come farebbero in un osso sesamoide come la rotula. Negli uccelli, l’ossificazione del tendine si verifica principalmente nell’arto posteriore, mentre nei dinosauri ornitischi, i tendini dei muscoli assiali ossificati formano un reticolo lungo le spine neurali ed ematiche della coda, presumibilmente per il sostegno.
Vedi anche
| Wikimedia Commons has media related to Tendons. |
- Aponeurosi
- Cartilagine
- Cordae tendineae
- Lista dei muscoli del corpo umano
- Guaina tendinea
- ^ Dorlands Medical Dictionary, pagina 602
- ^ Caldini, E. G.; Caldini, N.; De-Pasquale, V.; Strocchi, R.; Guizzardi, S.; Ruggeri, A.; Montes, G. S. (1990). “Distribuzione delle fibre del sistema elastico nel tendine della coda di ratto e nelle sue guaine associate”. Cellule Tessuti Organi. 139 (4): 341-348. doi:10.1159/000147022. PMID 1706129.
- ^ Grant, T. M.; Thompson, M. S.; Urban, J.; Yu, J. (2013). “Le fibre elastiche sono ampiamente distribuiti nel tendine e altamente localizzato intorno tenociti”. Giornale di Anatomia. 222 (6): 573-579. doi:10.1111/joa.12048. PMC 3666236. PMID 23587025.
- ^ Dorlands Medical Dictionary 2012.Page 1382
- ^ a b c Jozsa, L., e Kannus, P., Tendini umani: Anatomia, fisiologia e patologia. Human Kinetics: Champaign, IL, 1997.
- ^ Lin, T. W.; Cardenas, L.; Soslowsky, L. J. (2004). “Biomeccanica delle lesioni tendinee e la riparazione”. Journal of Biomechanics. 37 (6): 865-877. doi:10.1016/j.jbiomech.2003.11.005. PMID 15111074.
- ^ Kjær, Michael (aprile 2004). “Ruolo della matrice extracellulare in adattamento del tendine e muscolo scheletrico al carico meccanico”. Recensioni fisiologiche. 84 (2): 649-698. doi:10.1152/physrev.00031.2003. ISSN 0031-9333. PMID 15044685.
- ^ Taye, Nandaraj; Karoulias, Stylianos Z.; Hubmacher, Dirk (gennaio 2020). “L'”altro” 15-40%: Il ruolo delle proteine della matrice extracellulare non collagene e collageni minori nel tendine”. Journal of Orthopaedic Research. 38 (1): 23-35. doi:10.1002/jor.24440. ISSN 0736-0266. PMC 6917864. PMID 31410892.
- ^ Fukuta, S.; Oyama, M.; Kavalkovich, K.; Fu, F. H.; Niyibizi, C. (1998). “Identificazione dei tipi II, IX e X collageni al sito di inserzione del tendine d’achille bovino”. Matrice Biologia. 17 (1): 65-73. doi:10.1016/S0945-053X(98)90125-1. PMID 9628253.
- ^ Fratzl, P. (2009). “Cellulosa e collagene: dalle fibre ai tessuti”. Current Opinion in Colloid & Scienza dell’interfaccia. 8 (1): 32-39. doi:10.1016/S1359-0294(03)00011-6.
- ^ Zhang, G. E., Y.; Chervoneva, I.; Robinson, P. S.; Beason, D. P.; Carine, E. T.; Soslowsky, L. J.; Iozzo, R. V.; Birk, D. E. (2006). “La decorina regola l’assemblaggio delle fibrille di collagene e l’acquisizione delle proprietà biomeccaniche durante lo sviluppo del tendine”. Journal of Cellular Biochemistry. 98 (6): 1436-1449. doi:10.1002/jcb.20776. PMID 16518859. S2CID 39384363.CS1 maint: multiple names: authors list (link)
- ^ Raspanti, M.; Congiu, T.; Guizzardi, S. (2002). “Aspetti strutturali della matrice extracellulare del tendine: uno studio di forza atomica e microscopia elettronica a scansione”. Archivi di Istologia e Citologia. 65 (1): 37-43. doi:10.1679/aohc.65.37. PMID 12002609.
- ^ Scott, J. E. O., C. R.; Hughes, E. W. (1981). “Proteoglicano-collagene accordi in sviluppo tendine coda di ratto. Un’indagine microscopica elettronica e biochimica”. Biochemical Journal. 195 (3): 573-581. doi:10.1042/bj1950573. PMC 1162928. PMID 6459082.CS1 maint: multiple names: authors list (link)
- ^ Scott, J. E. (2003). “Elasticità in matrice extracellulare ‘moduli di forma’ del tendine, cartilagine, ecc Un modello di proteoglicano-filamento scorrevole”. Giornale di Fisiologia. 553 (2): 335-343. doi:10.1113/jphysiol.2003.050179. PMC 2343561. PMID 12923209.
- ^ McNeilly, C. M.; Banes, A. J.; Benjamin, M.; Ralphs, J. R. (1996). “Le cellule del tendine in vivo formano una rete tridimensionale di processi cellulari collegati da gap junctions”. Giornale di Anatomia. 189 (Pt 3): 593-600. PMC 1167702. PMID 8982835.
- ^ “Avere un tendine d’Achille corto può essere il tallone d’Achille di un atleta”. Recuperato 2007-10-26.
- ^ Young, Michael. “Una revisione sul riallineamento posturale e le sue componenti muscolari e neurali” (PDF).
- ^ Thorpe C.T., Birch H.L., Clegg P.D., Screen H.R.C. (2013). Il ruolo della matrice non collagene nella funzione del tendine. Int J ExpPathol. 94;4: 248-59.
- ^ Hulmes, D. J. S. (2002). “Costruire molecole di collagene, fibrille e strutture soprafibrillari”. Journal of Structural Biology. 137 (1-2): 2-10. doi:10.1006/jsbi.2002.4450. PMID 12064927.
- ^ Silver, F. H.; Freeman, J. W.; Seehra, G. P. (2003). “Collagene auto-assemblaggio e lo sviluppo delle proprietà meccaniche del tendine”. Journal of Biomechanics. 36 (10): 1529-1553. doi:10.1016/S0021-9290(03)00135-0. PMID 14499302.
- ^ Ker, R. F. (2002). “Le implicazioni della qualità di fatica adattabile dei tendini per la loro costruzione, riparazione e funzione”. Biochimica comparata e fisiologia A. 133 (4): 987-1000. doi:10.1016/S1095-6433(02)00171-X. PMID 12485688.
- ^ Cribb, A. M.; Scott, J.E. (1995). In Tendon response to tensile-stress – an ultrastructural investigation of collagen – proteoglycan interactions in stressed tendon,1995; Cambridge Univ Press.pp 423-428.
- ^ Screen H.R., Lee D.A., Bader D.L., Shelton J.C. (2004). “Un’indagine sugli effetti della struttura gerarchica dei fascicoli del tendine sulle proprietà micromeccaniche”. Proc Inst Mech Eng H. 218 (2): 109-119. doi:10.1243/095441104322984004. PMID 15116898. S2CID 46256718.CS1 maint: multiple names: authors list (link)
- ^ Puxkandl, R.; Zizak, I.; Paris, O.; Keckes, J.; Tesch, W.; Bernstorff, S.; Purslow, P.; Fratzl, P. (2002). “Proprietà viscoelastiche del collagene: indagini con radiazioni di sincrotrone e modello strutturale”. Philosophical Transactions of the Royal Society B. 357 (1418): 191-197. doi:10.1098/rstb.2001.1033. PMC 1692933. PMID 11911776.
- ^ Gupta H.S., Seto J., Krauss S., Boesecke P.& Schermo H.R.C. (2010). In situ analisi multilivello dei meccanismi di deformazione viscoelastica nel collagene del tendine. J. Struct. Biol. 169(2):183-191.
- ^ Thorpe C.T; Udeze C.P; Birch H.L.; Clegg P.D.; Screen H.R.C. (2012). “Specializzazione delle proprietà meccaniche del tendine risulta da differenze inter-fascicolari”. Journal of the Royal Society Interface. 9 (76): 3108-3117. doi:10.1098/rsif.2012.0362. PMC 3479922. PMID 22764132.
- ^ Thorpe C.T.; Klemt C; Riley G.P.; Birch H.L.; Clegg P.D.; Screen H.R.C. (2013). “Sottostrutture elicoidali in tendini di accumulo di energia forniscono un possibile meccanismo per l’accumulo e il ritorno efficiente di energia”. Acta Biomater. 9 (8): 7948-56. doi:10.1016/j.actbio.2013.05.004. PMID 23669621.
- ^ Gatt R, Vella Wood M, Gatt A, Zarb F, Formosa C, Azzopardi KM, Casha A, Agius TP, Schembri-Wismayer P, Attard L, Chockalingam N, Grima JN (2015). “Rapporti di Poisson negativi nei tendini: Una risposta meccanica inaspettata”. Acta Biomater. 24: 201-208. doi:10.1016/j.actbio.2015.06.018. PMID 26102335.
- ^ Batson EL, Paramour RJ, Smith TJ, Birch HL, Patterson-Kane JC, Goodship AE. (2003). Equine Vet J. |volume=35 |issue=3 |pages=314-8.Are the material properties and matrix composition of equine flexor and extensor tendons determined by their functions?
- ^ ScreenH.R.C., Tanner, K.E. (2012). Struttura & Biomeccanica dei compositi biologici. In: Enciclopedia dei Compositi 2nd Ed. Nicolais & Borzacchiello.Pub. John Wiley & Sons, Inc. ISBN 978-0-470-12828-2 (pagine 2928-39)
- ^ Nakagawa, Y. (1989). “Effetto del disuso sulla struttura ultra del tendine d’Achille nei ratti”. European Journal of Applied Physiology. 59 (3): 239-242. doi:10.1007/bf02386194. PMID 2583169. S2CID 20626078.
- ^ Reeves, N. D. (2005). “Influenza di 90 giorni di micro-gravità simulata sulle proprietà meccaniche del tendine umano e l’effetto delle contromisure di riposo”. Journal of Applied Physiology. 98 (6): 2278-2286. doi:10.1152/japplphysiol.01266.2004. hdl:11379/25397. PMID 15705722.
- ^ a b Riley, G. (2004). “La patogenesi della tendinopatia. Una prospettiva molecolare” (PDF). Reumatologia. 43 (2): 131-142. doi:10.1093/rheumatology/keg448. PMID 12867575.
- ^ a b c d Sharma, P. M., N. (2006). “Biologia delle lesioni tendinee: guarigione, modellazione e rimodellamento”. Journal of Musculoskeletal and Neuronal Interactions. 6 (2): 181-190. PMID 16849830.CS1 maint: multiple names: authors list (link)
- ^ a b c d Sharma, P.; Maffulli, N. (2005). “Lesione del tendine e tendinopatia: guarigione e riparazione”. Journal of Bone and Joint Surgery. Volume americano. 87A (1): 187-202. doi:10.2106/JBJS.D.01850. PMID 15634833. S2CID 1111422.
- ^ a b c d e f Wang, J. H. C. (2006). “Mechanobiology del tendine”. Giornale di Biomeccanica. 39 (9): 1563-1582. doi:10.1016/j.jbiomech.2005.05.011. PMID 16000201.
- ^ Riley, G. P.; Curry, V.; DeGroot, J.; van El, B.; Verzijl, N.; Hazleman, B. L.; Banca, R. A. (2002). “Attività delle metalloproteinasi di matrice e la loro relazione con il rimodellamento del collagene nella patologia del tendine”. Biologia della matrice. 21 (2): 185-195. doi:10.1016/S0945-053X(01)00196-2. PMID 11852234.
- ^ Moulin, V.; Tam, B. Y. Y.; Castilloux, G.; Auger, F. A.; O’Connor-McCourt, M. D.; Philip, A.; Germain, L. (2001). “I fibroblasti cutanei umani fetali e adulti mostrano differenze intrinseche nella capacità contrattile”. Journal of Cellular Physiology. 188 (2): 211-222. doi:10.1002/jcp.1110. PMID 11424088. S2CID 22026692.
- ^ Boyer, M. I. W., J. T.; Lou, J.; Manske, P. R.; Gelberman, R. H.; Cai, S. R. (2001). “Variazione quantitativa nell’espressione dell’mRNA del fattore di crescita endoteliale vascolare durante la guarigione precoce del tendine del flessore: un’indagine in un modello canino”. Journal of Orthopaedic Research. 19 (5): 869-872. doi:10.1016/S0736-0266(01)00017-1. PMID 11562135. S2CID 20903366.CS1 maint: multiple names: authors list (link)
- ^ Astrom, M.; Rausing, A. (1995). “Tendinopatia cronica di Achille – Un sondaggio dei risultati chirurgici e istopatologici”. Ortopedia clinica e ricerca correlata. 316 (316): 151-164. doi:10.1097/00003086-199507000-00021. PMID 7634699. S2CID 25486134.
- ^ Berge, James C. Vanden; Storer, Robert W. (1995). “Ossificazione intratendinea negli uccelli: Una revisione”. Giornale di Morfologia. 226 (1): 47-77. doi:10.1002/jmor.1052260105. PMID 29865323. S2CID 46926646.
- ^ Organ, Chris L. (2006). “Biomeccanica dei tendini ossificati in dinosauri ornitopodi”. Paleobiologia. 32 (4): 652-665. doi:10.1666/05039.1. S2CID 86568665.