
私たちは主に顔を合わせてお互いを認識し、コミュニケーションをとっています。 個々の顔の認識は、人間の顔が非常に多様であるからこそ可能なのである。 一卵性双生児は、両親から同じ遺伝子を受け継ぐため、遺伝子型が同じであり、顔が非常によく似ていることから、人を認識するためのさまざまな顔の特徴が受け継がれていることがわかります。 つまり、両親から受け継いだ遺伝子変異の特定の組み合わせによって、そのほとんどが決定されるのです。 遺伝子変異とは、ある遺伝子が、DNAのある位置で他の遺伝子と異なっていることを意味する。
鼻の形、後退した顎、「ハプスブルク唇」などの顔の特徴は、家族の中で世代を超えて受け継がれることが多い。 私たちの目的は、特定の顔立ちを決定する特定の遺伝子変異を特定することです。
私たちの出発点は、市販のハイテクカメラで撮影した3次元の顔画像の大規模なコレクションでした。 これらの画像は、いくつかの操作の後、顔の表面上の3万点として各顔を定義し、事実上、顔の3次元マップを提供してくれました。 異なる顔の特徴を比較するために、すべての顔画像を互いに重ね合わせました。 これは、鼻先や目尻、口元などのランドマークとなる点を基準に、画像を重ね合わせることで行います。 この方法は、顔や双子の研究のパイオニアであるフランシス・ガルトンが150年前に用いた方法とよく似ていますが、現在では高度なコンピューターツールやハイテクカメラにより、すべての画像を互いに重ね合わせることができる範囲が非常に広くなっています。
私たちは3つのソースからボランティアを使用しました。a) 私たちの非常によく特徴付けられたPoBI (People of the British Isles) 研究からの1832人のボランティア、b) TwinsUKコホートからの1567人のユニークな双子、ほぼ同じ数の一卵性双生児、c) 主に中国の東アジア人の画像33人です。 一卵性双生児は、同じ遺伝的変異(DNA配列)を持っています。 その結果、非常によく似た顔立ちになるのです。 顔の特徴は、3次元地図の丘や谷のように、顔の点の関連したグループによって定義されます。 この位置がどの程度異なるかは、この顔の点の位置に対する非遺伝的な環境的影響の指標となる。 一方、非一卵性双生児である2人は、顔の特徴の少なくとも一部を決定する遺伝的変異が異なる場合があります。 そのため、一卵性双生児の片方の顔画像上の同じ点の位置は、もう片方の双子の画像上の対応する点の位置と、一卵性双生児の場合ほど近くならない傾向がある。 一卵性双生児よりも非一卵性双生児の方が、どの程度ポイントが離れているかは、このポイントに対する遺伝的影響の指標となり、遺伝学者はこれを遺伝率と呼んでいる。
この重み付けの効果は、異なる遺伝率を持つ顔の輪郭上の点の頻度をプロットした図1に見ることができます。 任意の位置の遺伝性の程度は、測定値が一卵性双生児のペアで常にまったく同じであるが、非一卵性双生児では異なる場合の1から、一卵性双生児間の差が非一卵性双生児間の差と同じであり、事実上すべて
非遺伝的で主に環境的に決定する場合の0までである。 赤の列は重み付けした値、青の列は元の値、紫の列はオーバーラップした値である。 赤のプロファイルは青よりも明らかに平均的に高く、幅が狭く、重み付けの有益な効果を示している。
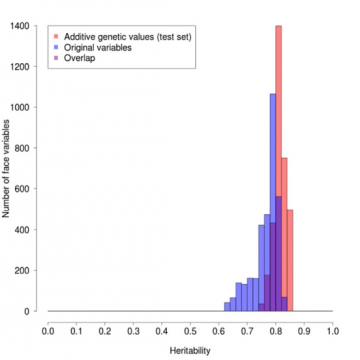
図1:オリジナルと遺伝率重み付け値のプロファイル遺伝率の比較
次の課題は、関連する点のグループに基づいて、遺伝分析に使用する顔の特徴を定義することである。 これには、重み付けされていないポイントを用いて得られた特徴よりも、全体的に遺伝性が高いと思われる特徴を与えるという仮定で、遺伝率で重み付けされたポイントを用いる。 重み付けされたポイントは、統計学者がPCA(Principal Components Analysis)と呼ぶ分析に使用されました。 これは、データから最も変動しやすい特徴を抽出する方法である。 各 PCA (顔画像のような複雑なデータでは 50 以上) は、それ自体が顔の特徴の測定値であり、目と目の間の距離に似ていますが、ポイントのグループに基づくさまざまな測定値を効果的に 1 つの値に結合するものです。 私たちは、PoBIのボランティアで画像がある約1500人、およびTwinsUKのボランティアで画像がある同数の人について、それぞれ約50万件のDNAベースの遺伝情報を持っています。
私たちの遺伝子解析のアプローチは、顔の特徴の違いは、身長のような定量的な尺度ではなく、個別の識別可能な特徴として解析されるべきであるという考えに基づいています。 身長だけでなく、目と目の間の距離や顔の縦横比など、顔の量的な特徴だけでは、その人を認識することはできない。 そこで、PCAで得られた各数値の極端な上位10%または下位10%に位置する個体に着目し、50万個の遺伝子マーカーのうち1つでも多く共有しているかどうかを問うことで、この問題に対処しているのです。 People of the British Islesのボランティアを用いて、50万個の遺伝子変異の中から、極端な個体とそうでない個体の差の有意性とこの差の大きさに基づいて、さらなる分析のための候補を選び出しました。 次に、これらの候補の影響のどれかが、約1500人のTwinsUKボランティアで再現されるかどうかを尋ねました。 このようにして、我々は比較的大きな効果を持つ3つの特異的で再現性のある遺伝子変異を特定しました。2つは顔のプロフィールの特徴について、1つは目の周りの領域についてです。 これらの3つのバリアントは、それぞれ同じ重要な位置に異なるDNA配列を持つパートナー
を持ち、それぞれのケースで正の相関を持つバリアントの集団頻度は約10%であり、パートナーの方が約90%と高い頻度を持っています。 私たちは、関連するバリアントをa、そのパートナーをAと呼び、遺伝子が対になっているように、これらのバリアントの組み合わせは、aa、Aa、AAの3種類があります。
PCDH15という遺伝子で見つかったこれらのバリアントの最初のものは、バリアントのコピーを両方持っている英国女性(aa)では、バリアントのコピーを一つ(Aa)または全く(AA)だけ持っている女性に比べて、図2Cに示した女性の特徴を持つ確率が7倍以上増加しました。 この変異体は、英国と東アジアの女性ボランティアの間で異なる特徴とも関連しています。 中国人の顔の平均である図2Aと、PoBIの個体のうちより中国人らしいグループの図2Bの、鼻と上唇の端が上向きで顎が後退していることに注目し、図2Cと対比してみてください。 PCDH15遺伝子の産物は、発育中のマウスの鼻の嗅覚細胞や軟骨に見られることから、
我々がヒトで発見した変種の鼻への影響の可能性と一致する。
2番目の変種はMBTPS1という遺伝子で、図3に示した顔の違いに関連している。 この違いは女性で観察され、顔の特徴的なサブセットはバリアントの両方のコピー(aa)を持っていました。 この場合、上極端な表現型(図3A)に関連する遺伝子変異は、アフリカミドリザル、マカク、オリーブヒヒに(おそらくaaとして)存在し、そのパートナーである共通変異は、オランウータン、ゴリラ、チンパンジー、マーモセットに(おそらくAAとして)存在しており、この変異の違いが、これらの霊長類のグループの間の顔の違いに関わっている可能性が示唆された。
3つ目のバリアントは、TMEM163という遺伝子で、図4に示すように、男女ともに目の違いに関連している。 この遺伝子の欠損型はムコリピドーシスIV型と呼ばれる、顔面、特にまぶたの周りの異常を時々伴う病気に関与している可能性がある。 我々の研究では、図4Aに示すように、上方の極端に関連するのは、変異体(aa)の両方のコピーを持つ個体の部分集合である。 目の幅と目の高さ(眉毛の下からまぶたの上まで)は、どちらも下の極値より上の極値で大きいことに注意。
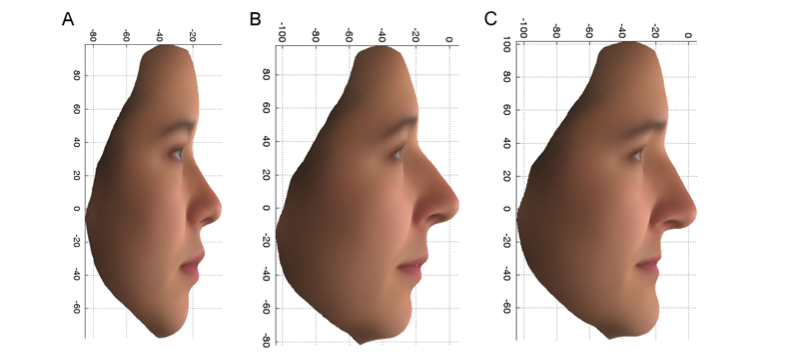
図2:PC2プロファイル。 東アジア人女性14名(A)とPoBI女性の上位10%(東アジア寄り)(B)と下位10%(ヨーロッパ寄り)(C)の平均顔、元の変数を使用した場合。
我々が特定の顔の特徴と関連付けることができた3つの遺伝子変異のそれぞれは、その変異のコピーを両方(aa)持っている英国のボランティアでは、その変異のコピーを1つ(Aa)または全く(AA)持っていない人と比べて、特定の特徴を持つ確率を
7倍以上増加させる。

Figure 3: PC7 profile: 元の変数を使用した、女性の顔の平均プロファイル、上部バリアント関連10%(A)および下部10%(C)の極値および全体の平均(B).

図4:PC1目。
これらの遺伝的変異を見つけることの成功は、双子のデータに基づいて高い遺伝率を持つ顔の特徴を識別する能力と、遺伝的変異の関連を研究するための極値の選択に大きく依存します。 今後、今回紹介したようなアプローチで、人間の顔の特徴に対する、より具体的で比較的大きな遺伝子変異の影響が多数発見される可能性があると思われます。 これは、遺伝的変異が人間の顔立ちの異常な多様性を決定する分子メカニズムを解明するための道筋を示すものである。