
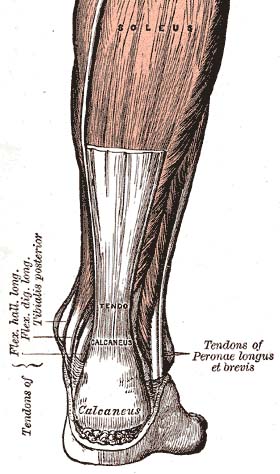

tendo
D013710
H3.H
Latin>
MeSHDetail Detail
Anatomical terminology
A tendon or sinew is a tough band of fibrous connective tissue that connects muscle to bone and are capable tostand tension.
Tendons are similar to ligament; both are made of collagen.The tended in a long termin. 靭帯は骨と骨をつなぎ、腱は筋肉と骨をつなぎます。
構造
組織学的に、腱は密な規則正しい結合組織からなります。 腱の主な細胞成分は、腱細胞という特殊な線維芽細胞である。 腱細胞は腱の細胞外マトリックスを合成し、コラーゲン繊維が密に詰まった豊富なマトリックスを形成する。 コラーゲン線維は互いに平行で、束状に組織化されている。 個々の腱束は、細いコラーゲン線維と弾性線維を含む繊細な緩い結合組織である腱内膜によって結合されている。 筋膜の集団は、密な不規則結合組織の鞘であるエピテノンに囲まれている。 腱全体は筋膜に包まれています。 筋膜と腱組織の間は、脂肪性乳糜爛組織であるパラテノンで満たされている。 正常な健康な腱は、シャーペイ繊維によって骨に固定されている。
細胞外マトリックス
正常な腱の乾燥質量は、その総質量の30~45%を占めている。
- 60-85%コラーゲン
- 60-80%コラーゲンI
- 0-10%コラーゲンIII
- 2%コラーゲンIV
- 少量のコラーゲンV、VI、その他
- 15-40%コラーゲン以外の細胞外マトリックスのコンポーネント、である。
- 3% 軟骨オリゴマー・マトリックス・タンパク、
- 1-2% エラスチン、
- 1-5% プロテオグリカン、
- 0.2%無機成分(銅、マンガン、カルシウムなど)。
腱のコラーゲンの大部分はコラーゲンIですが、腱の適切な発達と機能において重要な役割を果たすマイナーコラーゲンも多く存在しています。 軟骨部には II 型コラーゲン、血管壁のレチクリン繊維には III 型コラーゲン、毛細血管の基底膜には IX 型コラーゲン、血管壁には V 型コラーゲン、骨との境界付近の鉱化線維軟骨には X 型コラーゲンなどが含まれます。 細胞から分泌された後、プロコラーゲンN-およびC-プロテアーゼによって切断され、トロポコラーゲン分子は自然に不溶性の線維に集合する。 コラーゲン分子は長さ約300 nm、幅1-2 nmで、形成されるフィブリルの直径は50-500 nmに及ぶ。 腱では、フィブリルはさらに集合して、長さ約10mm、直径50-300μmの束を形成し、最終的に直径100-500μmの腱繊維になる。
腱のコラーゲンは、デコリンや、腱の圧縮領域ではアグリカンを含むプロテオグリカン(グリコサミノグリカン基にタンパク質が結合した化合物で、特に結合組織に存在)成分で結合されており、特定の位置でコラーゲン線維に結合する能力がある。 プロテオグリカンはコラーゲン線維に絡みつき、そのグリコサミノグリカン(GAG)側鎖は線維の表面と多重に相互作用しており、プロテオグリカンが線維の結合に構造的に重要であることが示されている。 腱の主要なGAG成分はデルマタン硫酸とコンドロイチン硫酸であり、これらはコラーゲンと会合し、腱の発達過程で線維の形成過程に関与している。 デルマタン硫酸は線維間の会合形成に関与し、コンドロイチン硫酸は線維間の容積を占め、線維を分離し、変形に耐えるのに役立つと考えられている。 デコリンのデルマタン硫酸側鎖は溶液中で凝集し、この挙動がコラーゲン線維の集合を助けると考えられている。 デコリン分子がコラーゲン線維に結合すると、そのデルマタン硫酸鎖が伸びて、別の線維に結合しているデコリン上の他のデルマタン硫酸鎖と会合し、線維間橋を作り、最終的に線維を平行に配列させることがある。 線維の束は繊維状に組織化され、その間に細長い腱細胞が密に詰まっている。 腱には、コラーゲンに関連した細胞突起の3次元ネットワークが存在する。 腱細胞はギャップ結合を通して互いに連絡を取り合い、このシグナル伝達により機械的負荷を感知して反応する能力を持つ。
腱内にはコラーゲン線維と平行に走る血管が見えることがあり、時折分岐した横吻合も見られる。
腱の内部には神経線維はないと考えられていますが、エピテノンとパラテノンには神経終末があり、腱と筋肉の接合部にはゴルジ腱器官が存在します。 腱の長さは、実際には、実際の筋肉の大きさと潜在的な筋肉の大きさに関する決定的な要因である。 たとえば、他のすべての関連する生物学的要因が同じである場合、腱が短く、上腕二頭筋が長い人は、腱が長く、筋肉が短い人よりも筋肉量の潜在的な可能性が高くなります。 ボディビルダーとして成功する人は、一般的に腱が短いと言われています。 腱の長さは遺伝的素因によって決定され、環境に応じて増加または減少することは示されていませんが、筋肉は外傷、使用上の不均衡、回復とストレッチの不足によって短くなる可能性があります。 RAT
機能
 腱の拡大図
腱の拡大図従来、腱は筋肉そのものと同様に筋肉と骨をつなぐ機構と考えられ、力を伝達する機能を担ってきました。 この接続により、腱は運動中に受動的に力を調節し、能動的な作業を行わずに追加の安定性を提供することができる。 しかし、過去20年以上にわたって、一部の腱の弾性特性やバネとしての機能に多くの研究がなされてきました。 すべての腱が同じ役割を果たすわけではありません。文字を書くときの指のように、主に手足の位置を決める腱(位置決め腱)もあれば、運動効率を上げるためにバネとして働く腱(エネルギー蓄積腱)もあります。 エネルギー貯蔵腱は、エネルギーを高効率で貯蔵・回収することができる。 例えば、人間の歩幅では、足首の関節を背屈させるときにアキレス腱が伸びる。 そして、歩行の終盤、足が底屈する(つま先を下に向ける)ときに、蓄えられた弾性エネルギーが放出される。 さらに、腱が伸びることで、筋肉は長さをあまり変えずに機能することができ、筋肉がより大きな力を生み出すことが可能になります。 コラーゲン線維は互いに平行で密に詰まっていますが、数マイクロメートルのスケールで平面的な起伏、すなわちクリンプにより波状の外観を示しています。 腱では、アミノ酸配列の特定位置にヒドロキシプロリンとプロリン残基がないため、コラーゲン線維はある程度の柔軟性を持ち、三重らせんの屈曲や内部ループなど他のコンフォーメーションの形成が可能となり、クリンプの発生に至る。 コラーゲン線維のクリンプにより、腱はある程度の柔軟性を持つと同時に、低い圧縮剛性を持つようになる。 さらに、腱は部分的に独立した多くの線維と束からなる多鎖構造であるため、一本の棒として振る舞うことはなく、この特性も柔軟性に寄与しています
腱のプロテオグリカン成分も機械特性に対して重要です。 コラーゲン線維によって腱が引張応力に耐えられるのに対して、プロテオグリカンによって圧縮応力に耐えることができるのです。 プロテオグリカンは、非常に親水性が高く、大量の水を吸収することができるため、膨潤率が高い。 プロテオグリカンは線維に非共有結合しているため、可逆的に結合と解離を繰り返し、線維間の橋が壊れたり、再構築されたりすることがある。 この過程は、線維が伸長し、張力下で直径が減少することに関与していると考えられる。 しかし、プロテオグリカンもまた、腱の引張特性に関与している可能性がある。 腱の構造は、事実上、繊維複合材料であり、一連の階層的なレベルとして構築されている。 各階層では、コラーゲン単位がコラーゲン架橋またはプロテオグリカンによって結合され、引張荷重に非常に強い構造を形成している。 コラーゲン線維だけの伸びやひずみは、同じ応力下での腱全体の伸びやひずみの合計よりもはるかに小さいことが示されており、プロテオグリカンに富むマトリックスも変形を受ける必要があり、マトリックスの硬化は高いひずみ速度で起こることが実証されています。 この非コラーゲン性マトリックスの変形は、腱の階層のすべてのレベルで発生し、このマトリックスの組織と構造を調節することによって、異なる腱に求められる異なる力学的特性を実現することができます。 エネルギー貯蔵型腱は、必要とされる高歪み特性を実現するために、筋膜間のスライドを多量に利用することが示されており、一方、位置制御型腱は、コラーゲン線維と線維間のスライドに大きく依存しています。 しかし、最近のデータから、エネルギーを蓄積する腱は、本質的にねじれた、またはらせん状の筋膜を含んでいる可能性があることが示唆されています。 軟組織
腱は粘弾性構造であり、弾性と粘性の両方の挙動を示す。 腱は伸ばされると、典型的な「軟部組織」としての挙動を示します。 力-伸び、または応力-ひずみ曲線は、クリンプ構造がまっすぐになり、コラーゲン繊維が整列するため、非常に低い剛性領域から始まり、腱の繊維の負のポアソン比を示唆します。 最近では、in vivo (MRI) および ex vivo (様々な死体腱組織の力学試験) で行われた試験により、健康な腱は非常に異方性が高く、長さ方向に2%まで、つまり通常の可動域内で伸ばしたときに、いくつかの平面で負のポアソン比 (auxetic) を示すことが分かっています。 この「つま先」領域を過ぎると、構造は著しく硬くなり、破損が始まるまで直線的な応力-ひずみ曲線を描くようになります。 腱の機械的特性は、腱の機能要件に適合しているため、大きく変化します。 エネルギーを蓄積する腱は、より簡単にエネルギーを蓄積できるよう、より弾性的、またはより硬くなる傾向があり、一方、より硬い位置決め腱は、動きをより細かく制御できるよう、やや粘弾性的、またはより弾性的になる傾向があります。 典型的なエネルギー貯蔵腱は、約12~15%のひずみと100~150MPaの応力で破損しますが、これより著しく伸びる腱もあります。例えば、馬の表在性趾屈筋は駆け足で20%以上伸びることがあります。 位置決め腱は、6~8%という低いひずみで破損することがありますが、700~1000MPaの弾性率を持つことがあります。
いくつかの研究により、腱は骨と同様に成長およびリモデリングのプロセスで機械的負荷の変化に反応することが証明されています。 特に、ラットのアキレス腱を廃用すると、腱を構成するコラーゲン繊維束の平均厚さが減少することが研究で示されました。 また、ヒトの場合、微小重力環境を模擬した実験では、安静運動をさせても、腱の硬さが著しく低下することがわかりました。 これらの効果は、寝たきりの患者の治療から、宇宙飛行士のより効果的なエクササイズの設計に至るまで、さまざまな分野に影響を及ぼしています。 そのため、骨折した腱の治癒過程は長く、痛みを伴います。 負傷から48時間以内に医師の診察を受けない場合、ほとんどの人が、負傷した場所のひどい腫れや痛み、灼熱感に悩まされます。
腱はマトリックスのターンオーバーが起こらず、腱細胞には修復能力がないと考えられていました。 しかし、人の一生を通じて、腱の腱細胞はマトリックス成分を活発に合成し、マトリックスメタロプロテアーゼ (MMP) などの酵素がマトリックスを分解することが明らかになって以来、腱はマトリックスのターンオーバーを起こさず、修復する能力がないと考えられてきました。 腱は、腱細胞およびその周囲の細胞外マトリックスによって制御されるプロセスで、損傷を治癒し回復することができます。
腱治癒の3つの主要段階は、炎症、修復または増殖、およびリモデリングで、さらに連結と成熟に分けることができます。 これらの段階は互いに重なり合うことがある。 第一段階では、好中球のような炎症細胞が赤血球とともに損傷部位に集められる。 単球とマクロファージは最初の24時間以内に採用され、損傷部位の壊死性物質の貪食が起こる。 血管作動性因子と走化性因子が放出された後、血管新生と腱細胞の増殖が開始される。 その後、腱細胞は傷害部位に移動し、コラーゲンIIIの合成を開始する。 数日後、修復または増殖の段階が始まる。 この段階では、腱細胞は損傷部位で大量のコラーゲンとプロテオグリカンの合成に関与し、GAGと水のレベルが高くなる。 約6週間後、リモデリング段階が始まる。 この段階の最初の部分は、受傷後約6週間から10週間続くコンソリデーションである。 この間、コラーゲンやGAGの合成は減少し、コラーゲンIの産生が増加して組織がより線維化し、線維が機械的ストレスの方向に整列するため、細胞数も減少する。 最終的な成熟段階は10週間後で、この間にコラーゲン線維の架橋が進み、組織が硬くなる。 徐々に、約1年かけて、組織は繊維状から瘢痕状に変化します。
Matrix Metalloproteinases (MMPs) は、腱損傷後の治癒過程におけるECMの分解とリモデリングに非常に重要な役割を担っています。 MMP-1、MMP-2、MMP-8、MMP-13、および MMP-14 を含む特定の MMP はコラゲナーゼ活性を有し、他の多くの酵素とは異なり、コラーゲン I 線維を分解することができることを意味している。 MMP-1によるコラーゲン線維の分解と変性コラーゲンの存在は、腱のECMを弱体化させ、別の断裂が発生する可能性を高めると考えられている要因である。 繰り返される機械的負荷や損傷に反応して、サイトカインが腱細胞から放出され、MMPの放出を誘導し、ECMの分解を引き起こし、損傷の再発や慢性腱鞘炎を引き起こす可能性がある
その他の様々な分子が腱の修復と再生に関与している。 腱の治癒中に有意にアップレギュレートされ活性化されることが示されている5つの成長因子があります:インスリン様成長因子1(IGF-I)、血小板由来成長因子(PDGF)、血管内皮成長因子(VEGF)、基本線維芽細胞成長因子(bFGF)、トランスフォーミング成長因子β(TGF-β)です。 これらの成長因子はすべて、治癒の過程で異なる役割を担っている。 IGF-1は炎症の第一段階でコラーゲンとプロテオグリカンの産生を増加させ、PDGFは損傷後の初期段階にも存在し、DNAの合成と腱細胞の増殖とともに他の成長因子の合成を促進します。 TGF-βの3つのアイソフォーム(TGF-β1、TGF-β2、TGF-β3)は、創傷治癒と瘢痕形成に関与することが知られています。 VEGFは血管新生を促進し、内皮細胞の増殖と移動を誘導することがよく知られており、VEGF mRNAはコラーゲンI mRNAとともに腱損傷部位で発現することが示されている。 骨形成タンパク質(BMP)はTGF-βスーパーファミリーのサブグループで、骨や軟骨の形成や組織の分化を誘導することができ、特にBMP-12は腱組織の形成と分化に影響を与え、線維形成を促進することが示されている。
治療に対する活動の影響
動物モデルにおいて、活動レベルという形での機械的負荷が腱損傷と治癒に対してどのような影響があるか、幅広い研究が行われた。 ストレッチは初期の炎症期には治癒を妨げる可能性がありますが、急性損傷から約1週間後に腱を制御して動かすと、腱細胞によるコラーゲンの合成が促進され、治癒した腱の引張強度と直径が増し、固定された腱よりも癒着が少なくなることが示されています。 また、慢性の腱損傷では、機械的負荷がコラーゲンの再調整とともに線維芽細胞の増殖とコラーゲン合成を刺激し、これらのすべてが修復とリモデリングを促進することが示されています。 運動や活動が腱の治癒を助けるという理論をさらに裏付けるように、損傷後の腱の固定はしばしば治癒にマイナスの影響を与えることが分かっています。 ウサギでは、固定されたコラーゲン束は引張強度の低下を示し、固定はまた腱の水分量、プロテオグリカン、コラーゲン架橋の減少をもたらす。
機械的力に対する腱細胞の反応の理由として、遺伝子発現、タンパク質合成、細胞の表現型を変え、最終的に腱構造の変化を引き起こすいくつかの機械伝達メカニズムが提案されてきた。 主な要因は、細胞外マトリックスの機械的変形であり、アクチン細胞骨格に影響を与え、その結果、細胞の形状、運動性、機能に影響を及ぼす可能性がある。 機械的な力は、焦点接着部位、インテグリン、細胞-細胞接合によって伝達されうる。 アクチン細胞骨格の変化は、インテグリンを活性化し、細胞とマトリックス間の「アウトサイドイン」と「インサイドアウト」シグナル伝達を媒介することができる。 また、細胞内シグナル伝達カスケードを誘導するGタンパク質も重要であると考えられ、イオンチャネルは伸展によって活性化され、カルシウム、ナトリウム、カリウムなどのイオンが細胞内に入ることができるようになります。 具体的な用途としては、縫い糸として使う、矢に羽を付ける(fletch 参照)、道具の刃をシャフトに括り付ける、などがあります。 また、サバイバル・ガイドでは、トラップや生活構造物などの丈夫な紐を作る材料として推奨されています。 このような用途に使用するためには、腱を特殊な方法で処理する必要があります。 イヌイットやその他の北極圏の人々は、彼らの生態系に適した他の繊維源がなかったため、あらゆる家庭用品の紐としてシニュー(腱)を利用してきました。 また、ユーラシア大陸の草原遊牧民やアメリカ先住民が好んで使った複合弓には、特定のシニューの弾性特性が生かされている。 最初の投石砲もシニューの弾性特性を利用したものでした。
シニューは、3 つの理由から優れた紐の材料になります。
料理への利用
主な記事: 天丼(食事)天丼(特に牛すじ)は、一部のアジア料理(飲茶や点心の店でよく出される)で食品として使用されている。 人気料理のひとつに、腱をニンニクでマリネしたsuan bao niu jinがある。 また、ベトナムの麺料理phở
臨床的意義
傷害
腱には多くの種類の傷害が存在する。 腱の損傷には様々な種類があり、使い過ぎによる腱の損傷もあります。 これらのタイプの損傷は、一般に腱の炎症と変性または弱化を引き起こし、最終的には腱の断裂につながる可能性があります。 腱障害は、腱の細胞外マトリックス(ECM)に関連する多くの要因によって引き起こされる可能性があり、その症状や病理組織はしばしば類似しているため、分類は困難でした。
腱障害の最初のカテゴリは傍脊椎炎で、これは腱と鞘の間にある傍脊椎シート、または傍脊椎板の炎症を指します。 腱鞘炎は、腱が細胞レベルで非炎症性の損傷を受けた状態を指します。 腱のコラーゲン、細胞、血管成分の損傷により劣化し、破断に至ることが知られています。 自然断裂を起こした腱を観察すると、コラーゲン線維が正しい平行配向でなかったり、長さや直径が均一でないこと、また丸みを帯びた腱細胞、その他の細胞の異常、血管の生着が認められます。 その他、断裂に至らない腱鞘炎でも、コラーゲン線維の変性、配向の乱れ、細化が見られ、線維間のグリコサミノグリカンの量も増加することが分かっています。 3つ目は、パラテノンの炎症と腱の変性がともに見られる、腱鞘炎を伴うパラテノン炎です。 最後に、腱炎であるが、これは腱の炎症と血管の破壊を伴う変性を指す。
腱障害は、年齢、体重、栄養などのいくつかの内因性要因によって引き起こされることがある。 外的要因としては、スポーツに関連することが多く、過度の力や負荷、不十分なトレーニング技術、環境条件などが挙げられます。
その他の動物
 Ossified tendon from an Edmontosaurus bone bed in Wyoming (Lance Formation)
Ossified tendon from an Edmontosaurus bone bed in Wyoming (Lance Formation)一部の生物、特に鳥や鳥竜類などの恐竜では腱が骨化することがあります。 この過程では、骨細胞が腱に浸潤し、膝蓋骨のような類骨で行われるように、骨を敷き詰めます。 鳥類では、腱の骨化は主に後肢で起こるが、鳥竜類では、軸筋腱が骨化し、尾部の神経棘と血棘に沿って格子状に形成されており、おそらく支持のためと思われる。
も参照
この記事は解剖学用語を使用しています。
Wikimedia Commons には腱に関するメディアもあります。 - 骨筋
- 軟骨
- 腱索
- 人体の筋肉一覧
- 腱鞘
- ^ Dorlands Medical Dictionary, 602ページ
- ^ Caldini, E. G.; Caldini, N.; De-Pasquale, V.; Strocchi, R.; Guizzardi, S.; Ruggeri, A.; Montes, G. S. (1990). 「ラット尾部腱とその関連鞘における弾性系繊維の分布”. 細胞 組織 臓器 139 (4): 341-348. doi:10.1159/000147022. PMID 1706129.
- ^ Grant, T. M.; Thompson, M. S.; Urban, J.; Yu, J. (2013). 「腱に広く分布し、腱細胞の周りに高度に局在している弾性線維”. ジャーナル・オブ・アナトミー。 222 (6): 573-579. doi:10.1111/joa.12048. PMC 3666236. PMID 23587025.
- ^ Dorlands Medical Dictionary 2012.Page 1382
- ^ a b c Jozsa, L., and Kannus, P.、Human Tendons: Anatomy, Physiology, and Pathology. Human Kinetics: Champaign, IL, 1997.
- ^ Lin, T. W.; Cardenas, L.; Soslowsky, L. J. (2004). 「腱の損傷と修復のバイオメカニクス」(Biomechanics of tendon injury and repair). ジャーナル・オブ・バイオメカニクス。 37 (6): 865-877. doi:10.1016/j.jbiomech.2003.11.005. PMID 15111074.
- ^ Kjær, Michael (April 2004). 「腱と骨格筋の機械的負荷への適応における細胞外マトリックスの役割”. Physiological Reviews. 84 (2): 649-698. doi:10.1152/physrev.00031.2003. issn 0031-9333. PMID 15044685.
- ^ Taye, Nandaraj; Karoulias, Stylianos Z.; Hubmacher, Dirk (January 2020). “その他 “の15~40%。 腱における非コラーゲン性細胞外マトリックスタンパク質とマイナーコラーゲンの役割”. Journal of Orthopaedic Research. 38 (1): 23-35. doi:10.1002/jor.24440. ISSN 0736-0266. PMC 6917864。 PMID 31410892.
- ^ Fukuta, S.; Oyama, M.; Kavalkovich, K.; Fu, F. H.; Niyibizi, C. (1998). 「ウシアキレス腱の挿入部におけるII型、IX型、X型コラーゲンの同定”. Matrix Biology. 17 (1): 65-73. doi:10.1016/S0945-053X(98)90125-1. PMID 9628253.
- ^ Fratzl, P. (2009). 「セルロースとコラーゲン:繊維から組織へ”. コロイド&界面科学における現在の意見。 8 (1): 32-39. doi:10.1016/S1359-0294(03)00011-6.
- ^ Zhang, G. E., Y.; Chervoneva, I.; Robinson, P. S.; Beason, D. P.; Carine, E. T.; Soslowsky, L. J.; Iozzo, R. V.; Birk, D. E. (2006). 「Decorin regulates assembly of collagen fibrils and acquisition of biomechanical properties during tendon development” (デコリンは腱の発達過程でコラーゲン線維の集合と生物力学的特性の獲得を制御する)。 ジャーナル・オブ・セルラー・バイオケミストリー。 98 (6): 1436-1449. doi:10.1002/jcb.20776. PMID 16518859. S2CID 39384363.CS1 maint: multiple names: authors list (link)
- ^ Raspanti, M.; Congiu, T.; Guizzardi, S. (2002). 「腱の細胞外マトリックスの構造的側面:原子間力顕微鏡と走査型電子顕微鏡の研究”. Archives of Histology and Cytology. 65 (1): 37-43. doi:10.1679/aohc.65.37. PMID 12002609.
- ^ Scott, J. E. O., C. R.; Hughes, E. W. (1981). 「ラット尾部腱におけるプロテオグリカン-コラーゲン配列。 An electron microscopical and biochemical investigation”. Biochemical Journal. 195 (3): 573-581. doi:10.1042/bj1950573. PMC 1162928. PMID 6459082.CS1 maint: multiple names: authors list (link)
- ^ Scott, J. E. (2003). “腱、軟骨などの細胞外マトリックス「形状モジュール」における弾性率。 A sliding proteoglycan-filament model”. ジャーナル オブ フィジオロジー。 553 (2): 335-343. doi:10.1113/jphysiol.2003.050179. PMC 2343561。 PMID 12923209.
- ^ McNeilly, C. M.; Banes, A. J.; Benjamin, M.; Ralphs, J. R. (1996). 「腱細胞は、ギャップジャンクションによってリンクされた細胞突起の3次元ネットワークを形成している」。 解剖学のジャーナル。 189 (Pt 3): 593-600. PMC 1167702. PMID 8982835.
- ^ “アキレス腱が短いことは、アスリートのアキレス腱かもしれない”. 2007-10-26 に掲載された。
- ^ Young, Michael. 「姿勢の再調整とその筋肉および神経コンポーネントに関するレビュー” (PDF).
- ^ ソープ C.T., バーチ H.L., クレッグ P.D., スクリーン H.R.C. (2013). 腱の機能における非コラーゲン性マトリックスの役割. Int J ExpPathol. 94;4: 248-59.
- ^ Hulmes, D. J. S. (2002). 「コラーゲン分子、フィブリル、および超フィブリル構造の構築”. ジャーナル・オブ・ストラクチュラル・バイオロジー。 137 (1-2): 2-10. doi:10.1006/jsbi.2002.4450. PMID 12064927.
- ^ Silver, F. H.; Freeman, J. W.; Seehra, G. P. (2003). 「コラーゲンの自己組織化と腱の力学的特性の発達”(Collagen self-assembly and the development of tendon mechanical properties). ジャーナル オブ バイオメカニクス。 36 (10): 1529-1553. doi:10.1016/S0021-9290(03)00135-0. PMID 14499302.
- ^ Ker, R. F. (2002). 「腱の適応性疲労品質がその構造、修復、機能に及ぼす影響”. PMID 12485688.
- ^ Cribb, A. M.; Scott, J.E. (1995). In Tendon response to tensile-stress – an ultrastructural investigation of collagen – proteoglycan interactions in stressed tendon,1995; Cambridge Univ Press.pp 423-428.
- ^ Screen H.R., Lee D.A., Bader D.L., Shelton J.C. (2004). 「腱の筋膜の階層構造がマイクロメカニカル特性に及ぼす影響に関する調査”. また、腱の筋膜の階層構造がマイクロメカニカル特性に及ぼす影響について検討した。 PMID 15116898. S2CID 46256718.CS1 maint: multiple names: authors list (link)
- ^ Puxkandl, R.; Zizak, I.; Paris, O.; Keckes, J.; Tesch, W.; Bernstorff, S.; Purslow, P.; Fratzl, P. (2002). 「コラーゲンの粘弾性特性:放射光による研究と構造モデル”. 王立協会Bの哲学的論文 357 (1418): 191-197. PMC 1692933。 PMID 11911776.
- ^ Gupta H.S., Seto J., Krauss S., Boesecke P.& Screen H.R.C. (2010). 腱コラーゲンの粘弾性変形メカニズムのin situマルチレベル解析。 J. Struct. Biol.169(2):183-191。
- ^ ソープ C.T; Udeze C.P; Birch H.L.; Clegg P.D.; Screen H.R.C. (2012). 「腱の機械的特性の特殊化は、筋膜間の差異から生じる”. 王立協会インターフェイスのジャーナル。 9 (76): 3108-3117. doi:10.1098/rsif.2012.0362. PMC 3479922。 PMID 22764132.
- ^ Thorpe C.T.; Klemt C; Riley G.P.; Birch H.L.; Clegg P.D.; Screen H.R.C. (2013). このような場合、「腱の螺旋副構造は、効率的なエネルギー貯蔵と還流のための可能なメカニズムを提供する」。 アクタバイオマター。 9 (8): 7948-56. doi:10.1016/j.actbio.2013.05.004. PMID 23669621.
- ^ Gatt R, Vella Wood M, Gatt A, Zarb F, Formosa C, Azzopardi KM, Casha A, Agius TP, Schembri-Wismayer P, Attard L, Chockalingam N, Grima JN (2015). “腱の負のポアソン比。 予期せぬ力学的応答”. Acta Biomater. 24: 201-208. doi:10.1016/j.actbio.2015.06.018. PMID 26102335.
- ^ Batson EL, Paramour RJ, Smith TJ, Birch HL, Patterson-Kane JC, Goodship AE. (2003). Equine Vet J. |volume=35 |issue=3 |pages=314-8.Are the material properties and matrix composition of equine flexor and extensor tendons determined by their functions?
- ^ ScreenH.R.C., Tanner, K.E. (2012). 構造 & 生物学的複合材料のバイオメカニクス. In: コンポジット百科事典 第2版. ニコライ & ボルザッキエーロ.パブ. ジョン・ワイリー&・サンズ,Inc. ISBN 978-0-470-12828-2 (pages 2928-39)
- ^ Nakagawa, Y. (1989). 「ラットにおけるアキレス腱の超構造に及ぼす廃用性の影響”. European ジャーナル・オブ・アプライド・フィジオロジー。 59 (3): 239-242. doi:10.1007/bf02386194. PMID 2583169. S2CID 20626078.
- ^ Reeves, N. D. (2005). “ヒト腱の力学的特性に対する90日間の模擬微小重力の影響と安静対策の効果”. Journal of Applied Physiology. 98 (6): 2278-2286. doi:10.1152/japplphysiol.01266.2004. hdl:11379/25397. PMID 15705722.
- ^ a b Riley, G. (2004). “The pathogenesis of tendinopathy. A molecular perspective” (PDF). Rheumatology. 43 (2): 131-142. doi:10.1093/rheumatology/keg448. PMID 12867575.
- ^ a b c d Sharma, P. M., N. (2006). 「腱損傷の生物学:治癒、モデリングとリモデリング”。 筋骨格系と神経系の相互作用のジャーナル。 6 (2): 181-190. PMID 16849830.CS1 maint: multiple names: authors list (link)
- ^ a b c d Sharma, P.; Maffulli, N. (2005). 「腱の損傷と腱障害:治癒と修復”。 骨と関節の手術のジャーナル。 アメリカのボリューム。 87A(1):187から202まで。 PMID 15634833。 S2CID 1111422.
- ^ a b c d e f Wang, J. H. C. (2006). 「腱のメカノバイオロジー”. ジャーナル・オブ・バイオメカニクス。 39 (9): 1563-1582. doi:10.1016/j.jbiomech.2005.05.011. PMID 16000201.
- ^ Riley, G. P.; Curry, V.; DeGroot, J.; van El, B.; Verzijl, N.; Hazleman, B. L.; Bank, R. A. (2002). 「Matrix metalloproteinase activities and their relationship with collagen remodelling in tendon pathology(マトリックスメタロプロテアーゼ活性と腱病理学におけるコラーゲン再構築との関係)”. Matrix Biology. 21 (2): 185-195. doi:10.1016/S0945-053X(01)00196-2. PMID 11852234.
- ^ Moulin, V.; Tam, B. Y. Y.; Castilloux, G.; Auger, F. A.; O’Connor-McCourt, M. D.; Philip, A.; Germain, L. (2001). 「このように、ヒトの皮膚線維芽細胞は、収縮能力において本質的な違いを示す。 細胞生理学のジャーナル。 188 (2): 211-222. doi:10.1002/jcp.1110. PMID 11424088. S2CID 22026692.
- ^ Boyer, M. I. W., J. T.; Lou, J.; Manske, P. R.; Gelberman, R. H.; Cai, S. R. (2001). 「腱の早期治癒における血管内皮増殖因子(VEGF)mRNAの発現量の変化:イヌモデルにおける検討」。 整形外科研究ジャーナル。 19 (5): 869-872. doi:10.1016/S0736-0266(01)00017-1. PMID 11562135. S2CID 20903366.CS1 maint: multiple names: authors list (link)
- ^ Astrom, M.; Rausing, A. (1995). 「慢性アキレス腱障害-外科的および病理組織学的所見の調査”. 臨床整形外科と関連研究。 316(316):151から164まで。 PMID 7634699. S2CID 25486134.
- ^ Berge, James C. Vanden; Storer, Robert W. (1995). 「鳥類の腱膜内骨化症。 A review”. ジャーナル・オブ・モルフォロジー。 226 (1): 47-77. doi:10.1002/jmor.1052260105. PMID 29865323. S2CID 46926646.
- ^ Organ, Chris L. (2006). 但し、”鳥脚類 “の骨化した腱のバイオメカニクスは、”鳥脚類 “の腱のバイオメカニクスと同じである。 Paleobiology. 32 (4): 652-665. s2cid 86568665.

