
Obiective de învățare
- Descrieți componentele implicate în contracția unui mușchi
- Explicați modul în care mușchii se contractă și se relaxează
- Descrieți modelul filamentului glisant al contracției musculare
Succesiunea de evenimente care duc la contracția unei fibre musculare individuale începe cu un semnal – neurotransmițătorul, ACh – de la neuronii motori care inervează acea fibră. Membrana locală a fibrei se va depolariza pe măsură ce intră ioni de sodiu încărcați pozitiv (Na+), declanșând un potențial de acțiune care se răspândește în restul membranei se va depolariza, inclusiv tubulii T. Acest lucru declanșează eliberarea de ioni de calciu (Ca++) din depozitul din reticulul sarcoplasmatic (SR). Ca++ inițiază apoi contracția, care este susținută de ATP (figura 1). Atâta timp cât ionii de Ca++ rămân în sarcoplasmă pentru a se lega de troponină, ceea ce menține „neprotejate” situsurile de legare a actinei, și atâta timp cât ATP este disponibil pentru a conduce ciclul punților încrucișate și tragerea firelor de actină de către miozină, fibra musculară va continua să se scurteze până la o limită anatomică.

Figura 1. Contracția unei fibre musculare. O punte încrucișată se formează între actină și capetele de miozină declanșând contracția. Atâta timp cât ionii de Ca++ rămân în sarcoplasmă pentru a se lega de troponină și atâta timp cât ATP este disponibil, fibra musculară va continua să se scurteze.
Contracția musculară se oprește, de obicei, atunci când se termină semnalizarea de la neuronii motori, care repolarizează sarcolema și tubulii T și închide canalele de calciu portate de tensiune din SR. Ionii de Ca++ sunt apoi pompați înapoi în SR, ceea ce face ca tropomiozina să resigileze (sau să acopere din nou) situsurile de legare de pe firele de actină. De asemenea, un mușchi se poate opri din contracție atunci când rămâne fără ATP și devine obosit (figura 2).

Figura 2. Relaxarea unei fibre musculare. Ionii de Ca++ sunt pompați înapoi în SR, ceea ce face ca tropomiozina să resigileze situsurile de legare de pe filamentele de actină. De asemenea, un mușchi se poate opri din contracție atunci când rămâne fără ATP și devine obosit.
Evenimentele moleculare de scurtare a fibrei musculare au loc în interiorul sarcomerilor fibrei (vezi figura 3). Contracția unei fibre musculare striate are loc pe măsură ce sarcomerele, dispuse liniar în cadrul miofibrilelor, se scurtează pe măsură ce capetele de miozină trag de filamentele de actină.
Regiunea în care filamentele groase și subțiri se suprapun are un aspect dens, deoarece există puțin spațiu între filamente. Această zonă în care filamentele subțiri și groase se suprapun este foarte importantă pentru contracția musculară, deoarece este locul unde începe mișcarea filamentelor. Filamentele subțiri, ancorate la capetele lor de către discurile Z, nu se extind complet în regiunea centrală care conține doar filamente groase, ancorate la baza lor într-un punct numit linia M. O miofibrilă este compusă din mai mulți sarcomere care se desfășoară de-a lungul lungimii sale; astfel, miofibrilele și celulele musculare se contractă pe măsură ce sarcomerele se contractă.
Modelul de contracție cu filamente glisante
Când este semnalat de un neuron motor, o fibră musculară scheletică se contractă pe măsură ce filamentele subțiri sunt trase și apoi alunecă pe lângă filamentele groase din cadrul sarcomerelor fibrei. Acest proces este cunoscut sub numele de modelul filamentului glisant al contracției musculare (figura 3). Glisarea poate avea loc numai atunci când situsurile de legare a miozinei pe filamentele de actină sunt expuse printr-o serie de etape care începe cu intrarea Ca++ în sarcoplasmă.

Figura 3. Modelul filamentului alunecător al contracției musculare. Atunci când un sarcomer se contractă, liniile Z se apropie una de alta, iar banda I devine mai mică. Banda A rămâne cu aceeași lățime. La contracția completă, filamentele subțire și gros se suprapun.
Tropomiozina este o proteină care se înfășoară în jurul lanțurilor filamentului de actină și acoperă situsurile de legare a miozinei pentru a împiedica legarea actinei de miozină. Tropomiozina se leagă de troponină pentru a forma un complex troponină-tropomiozină. Complexul troponină-tropomiozină împiedică „capetele” miozinei să se lege de situsurile active de pe microfilamentele de actină. Troponina are, de asemenea, un situs de legare pentru ionii Ca++.
Pentru a iniția contracția musculară, tropomiozina trebuie să expună situsul de legare a miozinei pe un filament de actină pentru a permite formarea punților încrucișate între microfilamentele de actină și miozină. Primul pas în procesul de contracție este ca Ca++ să se lege de troponină, astfel încât tropomiozina să poată aluneca departe de situsurile de legare de pe filamentele de actină. Acest lucru permite capetelor de miozină să se lege de aceste situsuri de legare expuse și să formeze punți încrucișate. Filamentele subțiri sunt apoi trase de capetele de miozină pentru a aluneca pe lângă filamentele groase spre centrul sarcomerului. Dar fiecare cap nu poate trage decât pe o distanță foarte scurtă înainte de a-și atinge limita și trebuie să fie „re-cocat” înainte de a putea trage din nou, o etapă care necesită ATP.
ATP și contracția musculară
Pentru ca filamentele subțiri să continue să alunece pe lângă filamentele groase în timpul contracției musculare, capetele de miozină trebuie să tragă actina la locurile de legare, să se desprindă, să se re-cocălească, să se atașeze la mai multe locuri de legare, să tragă, să se desprindă, să se re-cocălească, etc. Această mișcare repetată este cunoscută sub numele de ciclul punților încrucișate. Această mișcare a capetelor de miozină este similară cu cea a vâslelor atunci când un individ vâslește într-o barcă: Vâsla vâslelor (capetele de miozină) trag, sunt ridicate din apă (se detașează), repoziționate (se recockează) și apoi scufundate din nou pentru a trage (figura 4). Fiecare ciclu necesită energie, iar acțiunea capetelor de miozină din sarcomere care trag în mod repetat de filamentele subțiri necesită, de asemenea, energie, care este furnizată de ATP.
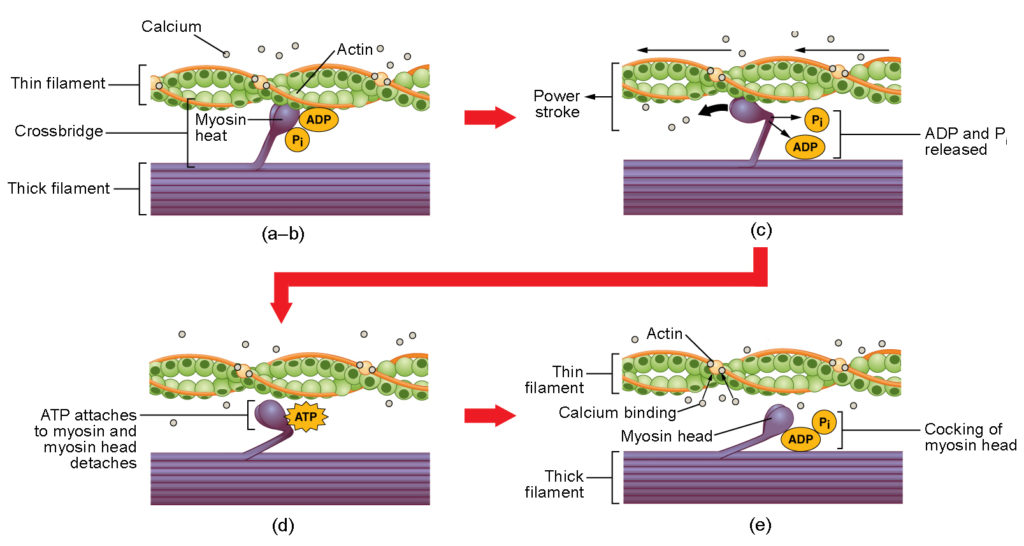
Figura 4. Contracția mușchiului scheletic. (a) Situl activ de pe actină este expus în timp ce calciul se leagă de troponină. (b) Capul miozinei este atras de actină, iar miozina se leagă de actină la situsul său de legare la actină, formând puntea încrucișată. (c) În timpul cursei de forță, fosfatul generat în ciclul anterior de contracție este eliberat. Acest lucru are ca rezultat pivotarea capului de miozină spre centrul sarcomerului, după care ADP-ul atașat și grupul fosfat sunt eliberate. (d) O nouă moleculă de ATP se atașează la capul de miozină, determinând detașarea punții încrucișate. (e) Capul de miozină hidrolizează ATP în ADP și fosfat, ceea ce readuce miozina în poziția armată.
Formarea punții încrucișate are loc atunci când capul de miozină se atașează de actină în timp ce adenozin difosfat (ADP) și fosfat anorganic (Pi) sunt încă legate de miozină (figura 4a,b). Pi este apoi eliberat, ceea ce face ca miozina să formeze o legătură mai puternică cu actina, după care capul de miozină se deplasează spre linia M, trăgând după sine actina. Pe măsură ce actina este trasă, filamentele se deplasează cu aproximativ 10 nm spre linia M. Această mișcare se numește cursa de putere, deoarece mișcarea filamentului subțire are loc în această etapă (figura 4c). În absența ATP, capul de miozină nu se va desprinde de actină.
O parte a capului de miozină se atașează la locul de legare de pe actină, dar capul are un alt loc de legare pentru ATP. Legarea ATP determină detașarea capului de miozină de actină (figura 4d). După ce acest lucru se întâmplă, ATP este transformat în ADP și Pi prin activitatea ATPază intrinsecă a miozinei. Energia eliberată în timpul hidrolizei ATP modifică unghiul capului miozinei într-o poziție înclinată (figura 4e). Capul miozinei este acum în poziție pentru mișcări ulterioare.
Când capul miozinei este înclinat, miozina se află într-o configurație cu energie ridicată. Această energie este consumată pe măsură ce capul de miozină se deplasează pe parcursul cursei de forță, iar la sfârșitul cursei de forță, capul de miozină se află într-o poziție cu energie scăzută. După lovitura de forță, ADP este eliberat; cu toate acestea, puntea încrucișată formată este încă în poziție, iar actina și miozina sunt legate între ele. Atâta timp cât ATP este disponibil, acesta se atașează ușor la miozină, ciclul punților încrucișate se poate repeta, iar contracția musculară poate continua.
Rețineți că fiecare filament gros de aproximativ 300 de molecule de miozină are mai multe capete de miozină, iar multe punți încrucișate se formează și se rup continuu în timpul contracției musculare. Înmulțiți acest lucru cu toți sarcomerii dintr-o miofibrilă, cu toate miofibrilele dintr-o fibră musculară și cu toate fibrele musculare dintr-un mușchi scheletic și puteți înțelege de ce este nevoie de atât de multă energie (ATP) pentru a menține mușchii scheletici în funcțiune. De fapt, pierderea de ATP este cea care duce la rigor mortis observată la scurt timp după ce cineva moare. Nefiind posibilă producerea de ATP în continuare, nu mai există ATP disponibil pentru ca capetele de miozină să se detașeze de locurile de legare a actinei, astfel încât punțile încrucișate rămân la locul lor, provocând rigiditatea mușchilor scheletici.
Surse de ATP
ATP furnizează energia necesară pentru ca contracția musculară să aibă loc. În plus față de rolul său direct în ciclul punților încrucișate, ATP furnizează, de asemenea, energie pentru pompele de transport activ Ca++ din SR. Contracția musculară nu are loc fără cantități suficiente de ATP. Cantitatea de ATP stocată în mușchi este foarte mică, suficientă doar pentru a alimenta contracții de câteva secunde. Pe măsură ce se descompune, ATP trebuie, prin urmare, să fie regenerat și înlocuit rapid pentru a permite o contracție susținută. Există trei mecanisme prin care ATP poate fi regenerat: metabolismul fosfatului de creatină, glicoliza anaerobă, fermentația și respirația aerobă.
Fosfatul de creatină este o moleculă care poate stoca energie în legăturile sale de fosfat. Într-un mușchi în repaus, excesul de ATP își transferă energia către creatină, producând ADP și fosfat de creatină. Aceasta acționează ca o rezervă de energie care poate fi utilizată pentru a crea rapid mai mult ATP. Atunci când mușchiul începe să se contracte și are nevoie de energie, fosfatul de creatină își transferă fosfatul înapoi la ADP pentru a forma ATP și creatină. Această reacție este catalizată de enzima creatin-kinază și are loc foarte rapid; astfel, ATP-ul derivat din creatin fosfat alimentează primele câteva secunde de contracție musculară. Cu toate acestea, fosfatul de creatină poate furniza energie doar pentru aproximativ 15 secunde, moment în care trebuie utilizată o altă sursă de energie (figura 5).

Figura 5. Metabolismul muscular. O parte din ATP este stocată într-un mușchi în repaus. În momentul în care începe contracția, acesta este consumat în câteva secunde. Mai mult ATP este generat de fosfatul de creatină timp de aproximativ 15 secunde.
Cum ATP-ul produs de fosfatul de creatină se epuizează, mușchii se orientează către glicoliză ca sursă de ATP. Glicoliza este un proces anaerob (care nu depinde de oxigen) care descompune glucoza (zahăr) pentru a produce ATP; cu toate acestea, glicoliza nu poate genera ATP la fel de repede ca și creatinfosfatul. Astfel, trecerea la glicoliză duce la o rată mai lentă de disponibilitate a ATP pentru mușchi. Zahărul utilizat în glicoliză poate fi furnizat de glucoza din sânge sau prin metabolizarea glicogenului care este stocat în mușchi. Descompunerea unei molecule de glucoză produce doi ATP și două molecule de acid piruvic, care pot fi utilizate în respirația aerobă sau, atunci când nivelul de oxigen este scăzut, transformate în acid lactic (figura 6).

Figura 6. Glicoliza și respirația aerobă. Fiecare moleculă de glucoză produce doi ATP și două molecule de acid piruvic, care pot fi utilizate în respirația aerobă sau transformate în acid lactic. Dacă oxigenul nu este disponibil, acidul piruvic este transformat în acid lactic, care poate contribui la oboseala musculară. Acest lucru apare în timpul exercițiilor fizice intense, când sunt necesare cantități mari de energie, dar oxigenul nu poate fi livrat suficient către mușchi.
Dacă oxigenul este disponibil, acidul piruvic este utilizat în respirația aerobă. Cu toate acestea, dacă oxigenul nu este disponibil, acidul piruvic este transformat în acid lactic, care poate contribui la oboseala musculară. Această conversie permite reciclarea enzimei NAD+ din NADH, care este necesară pentru ca glicoliza să continue. Acest lucru se întâmplă în timpul exercițiilor fizice intense, atunci când sunt necesare cantități mari de energie, dar oxigenul nu poate fi livrat suficient către mușchi. Glicoliza în sine nu poate fi susținută foarte mult timp (aproximativ 1 minut de activitate musculară), dar este utilă pentru a facilita scurte explozii de mare intensitate. Acest lucru se datorează faptului că glicoliza nu utilizează glucoza foarte eficient, producând un câștig net de doi ATP per moleculă de glucoză și un produs final de acid lactic, care poate contribui la oboseala musculară pe măsură ce se acumulează.
Respirația aerobă este descompunerea glucozei sau a altor nutrienți în prezența oxigenului (O2) pentru a produce dioxid de carbon, apă și ATP. Aproximativ 95 la sută din ATP necesar pentru mușchii în repaus sau moderat activi este furnizat de respirația aerobă, care are loc în mitocondrii. Intrările pentru respirația aerobă includ glucoza care circulă în fluxul sanguin, acidul piruvic și acizii grași. Respirația aerobă este mult mai eficientă decât glicoliza anaerobă, producând aproximativ 36 de ATP per moleculă de glucoză față de patru din glicoliză. Cu toate acestea, respirația aerobă nu poate fi susținută fără un aport constant de O2 în mușchiul scheletic și este mult mai lentă (figura 7). Pentru a compensa, mușchii stochează o cantitate mică de oxigen în exces în proteine numite mioglobină, permițând contracții musculare mai eficiente și mai puțină oboseală. Antrenamentul aerobic crește, de asemenea, eficiența sistemului circulator, astfel încât O2 poate fi furnizat mușchilor pentru perioade mai lungi de timp.

Figura 7. Respirația celulară. Respirația aerobă constă în descompunerea glucozei în prezența oxigenului (O2) pentru a produce dioxid de carbon, apă și ATP. Aproximativ 95 la sută din ATP necesar pentru mușchii în repaus sau moderat activi este furnizat de respirația aerobă, care are loc în mitocondrii.
Fatiga musculară apare atunci când un mușchi nu se mai poate contracta ca răspuns la semnalele sistemului nervos. Cauzele exacte ale oboselii musculare nu sunt pe deplin cunoscute, deși anumiți factori au fost corelați cu scăderea contracției musculare care apare în timpul oboselii. ATP este necesar pentru contracția normală a mușchilor, iar pe măsură ce rezervele de ATP sunt reduse, funcția musculară poate scădea. Acest lucru poate fi mai degrabă un factor în cazul unei producții musculare scurte, intense, decât în cazul unor eforturi susținute, de intensitate mai mică. Acumularea de acid lactic poate scădea pH-ul intracelular, afectând activitatea enzimatică și proteică. Dezechilibrele nivelurilor de Na+ și K+ ca urmare a depolarizării membranei pot perturba fluxul de Ca++ din SR. Perioadele lungi de exerciții fizice susținute pot deteriora SR și sarcolema, ducând la afectarea reglării Ca++.
Activitatea musculară intensă are ca rezultat o datorie de oxigen, care reprezintă cantitatea de oxigen necesară pentru a compensa ATP-ul produs fără oxigen în timpul contracției musculare. Oxigenul este necesar pentru a restabili nivelul ATP și al fosfatului de creatină, pentru a transforma acidul lactic în acid piruvic și, în ficat, pentru a transforma acidul lactic în glucoză sau glicogen. Alte sisteme utilizate în timpul exercițiilor fizice necesită, de asemenea, oxigen, iar toate aceste procese combinate au ca rezultat creșterea frecvenței respiratorii care apare după efort. Până când datoria de oxigen a fost satisfăcută, aportul de oxigen este ridicat, chiar și după ce exercițiul fizic a încetat.
Relaxarea unui mușchi scheletic
Relaxarea fibrelor musculare scheletice și, în cele din urmă, a mușchiului scheletic, începe cu neuronul motor, care încetează să mai elibereze semnalul său chimic, ACh, în sinapsa de la NMJ. Fibra musculară se va repolariza, ceea ce închide porțile din SR prin care era eliberat Ca++. Pompele acționate de ATP vor muta Ca++ din sarcoplasmă înapoi în SR. Acest lucru are ca rezultat „reasigurarea” situsurilor de legare a actinei pe filamentele subțiri. Fără capacitatea de a forma punți încrucișate între filamentele subțiri și cele groase, fibra musculară își pierde tensiunea și se relaxează.
Forța musculară
Numărul de fibre musculare scheletice dintr-un anumit mușchi este determinat genetic și nu se modifică. Forța musculară este direct legată de cantitatea de miofibrile și sarcomere din fiecare fibră. Factorii, cum ar fi hormonii și stresul (și steroizii anabolizanți artificiali), care acționează asupra mușchiului pot crește producția de sarcomere și miofibrile în interiorul fibrelor musculare, o schimbare numită hipertrofie, care are ca rezultat creșterea masei și volumului unui mușchi scheletic. De asemenea, diminuarea utilizării unui mușchi scheletic are ca rezultat atrofia, în care numărul de sarcomere și miofibrile dispare (dar nu și numărul de fibre musculare). Este obișnuit ca un membru în ghips să prezinte mușchi atrofiați atunci când ghipsul este îndepărtat, iar anumite boli, cum ar fi poliomielita, prezintă mușchi atrofiați.
Disfuncții ale sistemului muscular
Distrofia musculară Duchenne (DMD) este o slăbire progresivă a mușchilor scheletici. Este una dintre mai multe boli denumite colectiv „distrofie musculară”. DMD este cauzată de lipsa proteinei distrofină, care ajută filamentele subțiri ale miofibrilelor să se lege de sarcolemă. În lipsa unei cantități suficiente de distrofină, contracțiile musculare determină ruperea sarcolemei, provocând un aflux de Ca++, ceea ce duce la deteriorarea celulară și la degradarea fibrelor musculare. În timp, pe măsură ce leziunile musculare se acumulează, se pierde masa musculară și se dezvoltă deficiențe funcționale mai mari.
DMD este o tulburare ereditară cauzată de un cromozom X anormal. Afectează în principal bărbații și este de obicei diagnosticată în copilăria timpurie. DMD apare, de obicei, mai întâi ca o dificultate în ceea ce privește echilibrul și mișcarea, iar apoi progresează până la incapacitatea de a merge. Ea continuă să progreseze în sus în corp, de la extremitățile inferioare la partea superioară a corpului, unde afectează mușchii responsabili de respirație și circulație. În cele din urmă cauzează moartea din cauza insuficienței respiratorii, iar cei afectați nu trăiesc, de obicei, mai mult de 20 de ani.
Pentru că DMD este cauzată de o mutație în gena care codifică distrofina, s-a crezut că introducerea de mioblaste sănătoase în pacienți ar putea fi un tratament eficient. Mioblastele sunt celulele embrionare responsabile de dezvoltarea mușchilor și, în mod ideal, acestea ar fi purtătoare de gene sănătoase care ar putea produce distrofina necesară pentru contracția normală a mușchilor. Această abordare a fost în mare parte nereușită la om. O abordare recentă a implicat încercarea de a stimula producția de utrofină în mușchi, o proteină similară distrofinei care ar putea fi capabilă să preia rolul distrofinei și să prevină apariția daunelor celulare.
Întrebări de autoverificare
Realizați testul de mai jos pentru a vă verifica gradul de înțelegere a contracției și relaxării fibrelor musculare: