
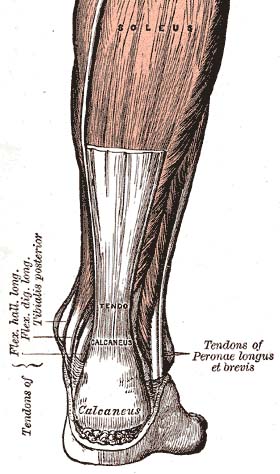

tendo
D013710
H3.03.00.0.00020
Anatomisk terminologi
En sena eller senor är ett tufft band av fibrös bindväv som förbinder muskeln med benet och som kan motstå spänningar.
Senor liknar ligament; båda består av kollagen. Ligament förbinder ett ben med ett annat, medan senor förbinder muskler med ben.
Struktur
Histologiskt sett består senor av tät regelbunden bindväv. Den huvudsakliga cellkomponenten i senor är specialiserade fibroblaster som kallas tenocyter. Tenocyterna syntetiserar senornas extracellulära matris, som är rikligt förekommande i tätt packade kollagenfibrer. Kollagenfibrerna är parallella med varandra och organiserade i fascikler. Enskilda fascikler är bundna av endotendineum, som är en känslig lös bindväv som innehåller tunna kollagenfibriller och elastiska fibrer. Grupper av fascikler avgränsas av epitenon, som är ett hölje av tät oregelbunden bindväv. Hela senan är omsluten av en fascia. Utrymmet mellan fascia och senvävnad är fyllt av paratenon, en fettrik areolär vävnad. Normala friska senor är förankrade i benet med Sharpeys fibrer.
Extracellulär matris
Den torra massan i normala senor, som utgör 30-45 % av deras totala massa, består av:
- 60-85% kollagen
- 60-80% kollagen I
- 0-10% kollagen III
- 2% kollagen IV
- måttliga mängder kollagen V, VI och andra
- 15-40% icke-kollagena komponenter i den extracellulära matrisen, inklusive:
- 3% brosk oligomeric matrix protein,
- 1-2% elastin,
- 1-5% proteoglykaner,
- 0.2 % oorganiska komponenter som koppar, mangan och kalcium.
Samtidigt som kollagen I utgör merparten av kollagenet i senan finns det många mindre kollagener som spelar viktiga roller för en korrekt utveckling och funktion av senan. Dessa inkluderar typ II-kollagen i broskzonerna, typ III-kollagen i retikulinfibrerna i kärlväggarna, typ IX-kollagen, typ IV-kollagen i kapillärernas basalmembran, typ V-kollagen i kärlväggarna och typ X-kollagen i det mineraliserade fibrbrosket nära gränssnittet mot benet.
Ultrastruktur och kollagensyntes
Kollagenfibrerna sammanfogas till makroaggregat. Efter utsöndring från cellen, som klyvs av procollagen N- och C-proteaser, samlas tropokollagenmolekylerna spontant till olösliga fibriller. En kollagenmolekyl är cirka 300 nm lång och 1-2 nm bred, och diametern på de fibriller som bildas kan variera mellan 50 och 500 nm. I senor samlas fibrillerna sedan ytterligare för att bilda fascikler, som är cirka 10 mm långa och har en diameter på 50-300 μm, och slutligen en senfiber med en diameter på 100-500 μm.
Kollagenet i senor hålls samman med proteoglykaner (en förening som består av ett protein bundet till glykosaminoglykangrupper och som framför allt förekommer i bindväv), bland annat decorin och, i komprimerade områden av senan, aggrekan, som kan binda sig till kollagenfibrillerna på specifika platser. Proteoglykanerna är sammanvävda med kollagenfibrillerna – deras glykosaminoglykan-sidokedjor (GAG) har flera interaktioner med fibrillernas yta – vilket visar att proteoglykanerna är strukturellt viktiga för sammankopplingen av fibrillerna. De viktigaste GAG-komponenterna i senan är dermatansulfat och kondroitinsulfat, som associerar med kollagen och är involverade i fibrillernas sammansättningsprocess under senans utveckling. Dermatansulfat anses vara ansvarigt för att bilda föreningar mellan fibriller, medan kondroitinsulfat anses vara mer involverat i att ta upp volymen mellan fibrillerna för att hålla dem separerade och hjälpa till att motstå deformation. Dekorins dermatansulfat-sidekedjor aggregerar i lösning, och detta beteende kan hjälpa till med sammansättningen av kollagenfibrillerna. När decorinmolekyler är bundna till en kollagenfibrill kan deras dermatansulfatkedjor sträcka ut sig och associera sig med andra dermatansulfatkedjor på decorin som är bundet till separata fibriller, vilket därför skapar interfibrillära broar och så småningom orsakar en parallell inriktning av fibrillerna.
Tenocyter
Tenocyterna producerar kollagenmolekylerna som aggregerar sig ända-till-änden och sida-till-sida för att producera kollagenfibriller. Fibrillbuntarna är organiserade för att bilda fibrer med de långsträckta tenocyterna tätt packade mellan dem. Det finns ett tredimensionellt nätverk av cellprocesser som är associerade med kollagenet i senan. Cellerna kommunicerar med varandra genom gap junctions, och denna signalering ger dem förmågan att upptäcka och reagera på mekanisk belastning.
Blodkärl kan visualiseras i endotendonet som löper parallellt med kollagenfibrerna, med enstaka förgrenade tvärgående anastomoser.
Den inre senans bulk anses inte innehålla några nervfibrer, men epitenon och paratenon innehåller nervändar, medan Golgi-senaorgan finns vid övergången mellan sena och muskel.
Senans längd varierar i alla huvudgrupper och från person till person. Senklängden är i praktiken den avgörande faktorn när det gäller faktisk och potentiell muskelstorlek. Om alla andra relevanta biologiska faktorer är lika kommer till exempel en man med kortare senor och längre bicepsmuskel att ha större potentiell muskelmassa än en man med längre senor och kortare muskel. Framgångsrika kroppsbyggare har i allmänhet kortare senor. Omvänt är det i sporter som kräver att atleterna utmärker sig i handlingar som löpning eller hoppning fördelaktigt att ha en längre än genomsnittlig akillessena och en kortare vadmuskel.
Senlängden bestäms av genetiska anlag och har inte visat sig vare sig öka eller minska som svar på miljön, till skillnad från muskler, som kan förkortas av trauma, obalans i användningen och bristande återhämtning och stretching. RAT
Funktioner

Traditionellt har senor betraktats som en mekanism genom vilken musklerna ansluter till benet samt musklerna i sig själva, som fungerar för att överföra krafter. Denna förbindelse gör det möjligt för senor att passivt modulera krafter under förflyttning, vilket ger ytterligare stabilitet utan aktivt arbete. Under de senaste två decennierna har dock mycket forskning fokuserat på de elastiska egenskaperna hos vissa senor och deras förmåga att fungera som fjädrar. Alla senor behöver inte ha samma funktionella roll, där vissa främst positionerar lemmar, t.ex. fingrarna när man skriver (positionssenor) och andra fungerar som fjädrar för att göra förflyttningen mer effektiv (energilagrande senor). Energilagrande senor kan lagra och återvinna energi med hög effektivitet. Under ett mänskligt steg sträcker sig t.ex. akillessenen när fotleden dorsalflexeras. Under den sista delen av steget, när foten plantarflexerar (pekar ner tårna), frigörs den lagrade elastiska energin. Eftersom senan sträcker sig kan muskeln dessutom fungera med mindre eller till och med ingen längdförändring, vilket gör att muskeln kan generera mer kraft.
Senans mekaniska egenskaper är beroende av kollagenfiberns diameter och orientering. Kollagenfibrillerna är parallella med varandra och tätt packade, men uppvisar ett vågliknande utseende på grund av plana böljor, eller krusningar, på en skala av flera mikrometer. I senor har kollagenfibrerna en viss flexibilitet på grund av avsaknaden av hydroxiprolin- och prolinrester på specifika ställen i aminosyrasekvensen, vilket gör det möjligt att bilda andra konformationer, t.ex. böjningar eller inre slingor i trippelhelixen, och resulterar i utvecklingen av krusningar. Krumningarna i kollagenfibrillerna gör det möjligt för senorna att ha en viss flexibilitet och en låg tryckstyvhet. Eftersom senan dessutom är en flersträngad struktur som består av många delvis oberoende fibriller och fascikler beter den sig inte som en enda stav, och denna egenskap bidrar också till dess flexibilitet.
Proteoglykankomponenterna i senor är också viktiga för de mekaniska egenskaperna. Medan kollagenfibrillerna gör att senorna kan motstå dragspänning, gör proteoglykanerna att de kan motstå tryckspänning. Dessa molekyler är mycket hydrofila, vilket innebär att de kan absorbera en stor mängd vatten och därför har en hög svällningsgrad. Eftersom de är icke-kovalent bundna till fibrillerna kan de reversibelt associera och disassociera så att broarna mellan fibrillerna kan brytas och återbildas. Denna process kan bidra till att fibrilen kan förlängas och minska i diameter under spänning. Proteoglykanerna kan dock också spela en roll för senans dragegenskaper. Senans struktur är i själva verket ett fiberkompositmaterial som är uppbyggt som en serie hierarkiska nivåer. På varje nivå i hierarkin är kollagenenheterna bundna till varandra av antingen kollagenkorslänkar eller proteoglykaner för att skapa en struktur som är mycket motståndskraftig mot dragbelastning. Det har visat sig att enbart kollagenfibrillernas förlängning och töjning är mycket lägre än den totala förlängningen och töjningen av hela senan vid samma belastning, vilket visar att den proteoglykanrika matrisen också måste deformeras, och att en förstyvning av matrisen sker vid höga töjningshastigheter. Denna deformation av den icke-kollagenhaltiga matrisen sker på alla nivåer i senans hierarki, och genom att modulera denna matris organisation och struktur kan man uppnå de olika mekaniska egenskaper som krävs för olika senor. Energilagrande senor har visat sig använda sig av betydande mängder glidning mellan fascikler för att möjliggöra de höga spänningsegenskaper som de kräver, medan positionella senor i högre grad förlitar sig på glidning mellan kollagenfibrer och fibriller. Nya uppgifter tyder dock på att energilagrande senor också kan innehålla fascikler som är vridna eller spiralformade – ett arrangemang som skulle vara mycket fördelaktigt för att ge det fjäderliknande beteende som krävs i dessa senor.
Mekanik
Senor är viskoelastiska strukturer, vilket innebär att de uppvisar både elastiskt och visköst beteende. När senor sträcks uppvisar de ett typiskt ”mjukvävnadsbeteende”. Kraft-sträcknings- eller spännings-deformationskurvan börjar med ett område med mycket låg styvhet, eftersom krympstrukturen rätar ut sig och kollagenfibrerna anpassar sig. Detta tyder på en negativ Poissonkvot i senans fibrer. På senare tid har tester in vivo (med hjälp av MRT) och ex vivo (genom mekanisk testning av olika typer av senvävnad från kadaver) visat att friska senor är mycket anisotropa och uppvisar en negativ Poissonkvot (auxetic) i vissa plan när de sträcks upp till 2 % i längden, dvs. inom det normala rörelseomfånget. Efter denna ”tå”-region blir strukturen betydligt styvare och har en linjär spännings-deformationskurva tills den börjar svikta. De mekaniska egenskaperna hos senor varierar kraftigt, eftersom de anpassas till senans funktionskrav. De energilagrande senorna tenderar att vara mer elastiska eller mindre styva, så att de lättare kan lagra energi, medan de styvare positionella senorna tenderar att vara lite mer viskoelastiska eller mindre elastiska, så att de kan ge en finare kontroll av rörelsen. En typisk energilagrande sena kommer att brista vid cirka 12-15 % töjning och en spänning i storleksordningen 100-150 MPa, även om vissa senor är betydligt mer töjbara än så, t.ex. den ytliga digitala böjaren hos hästar, som sträcker sig med mer än 20 % när de galopperar. Positionella senor kan brista vid så låga belastningar som 6-8 %, men kan ha moduler i storleksordningen 700-1000 MPa.
Flera studier har visat att senor reagerar på förändringar i mekanisk belastning med tillväxt- och remodelleringsprocesser, i likhet med ben. Särskilt en studie visade att avstängning av akillessenen hos råttor resulterade i en minskning av den genomsnittliga tjockleken på de kollagenfiberbuntar som ingår i senan. Hos människor visade ett experiment där människor utsattes för en simulerad mikrogravitationsmiljö att senans styvhet minskade avsevärt, även när försökspersonerna var tvungna att utföra återhämtningsövningar. Dessa effekter har konsekvenser på områden som sträcker sig från behandling av sängliggande patienter till utformning av effektivare övningar för astronauter.
Healing
Senorna i foten är mycket komplexa och invecklade. Därför är läkningsprocessen för en bruten sena lång och smärtsam. De flesta som inte får läkarvård inom de första 48 timmarna efter skadan kommer att drabbas av kraftig svullnad, smärta och en brännande känsla där skadan uppstod.
Man trodde att senor inte kunde genomgå matrisomsättning och att tenocyter inte var kapabla till reparation. Det har dock sedan dess visats att tenocyter i senan under hela livet aktivt syntetiserar matriskomponenter samt att enzymer som matrixmetalloproteinaser (MMP) kan bryta ner matrisen. Senor kan läka och återhämta sig från skador i en process som styrs av tenocyterna och deras omgivande extracellulära matris.
De tre huvudstadierna av senans läkning är inflammation, reparation eller proliferation och remodellering, som kan delas in ytterligare i konsolidering och mognad. Dessa stadier kan överlappa varandra. I det första stadiet rekryteras inflammatoriska celler som neutrofiler till skadeplatsen tillsammans med erytrocyter. Monocyter och makrofager rekryteras inom de första 24 timmarna, och fagocytos av nekrotiskt material på skadeplatsen uppstår. Efter frisättning av vasoaktiva och kemotaktiska faktorer inleds angiogenes och proliferation av tenocyter. Tenocyterna flyttar sedan in på platsen och börjar syntetisera kollagen III. Efter några dagar börjar reparations- eller proliferationsfasen. I detta skede är tenocyterna involverade i syntesen av stora mängder kollagen och proteoglykaner på skadeplatsen, och nivåerna av GAG och vatten är höga. Efter cirka sex veckor börjar remodelleringsfasen. Den första delen av detta skede är konsolideringen, som pågår mellan sex och tio veckor efter skadan. Under denna tid minskar syntesen av kollagen och GAGs, och cellulariteten minskar också eftersom vävnaden blir mer fibrös till följd av ökad produktion av kollagen I och fibrillerna blir inriktade i riktning mot den mekaniska påfrestningen. Det sista mognadsstadiet inträffar efter tio veckor, och under denna tid ökar tvärbindningen av kollagenfibrillerna, vilket gör att vävnaden blir styvare. Gradvis, under ungefär ett år, kommer vävnaden att övergå från fibrös till ärrliknande.
Matrixmetalloproteinaser (MMP) har en mycket viktig roll i nedbrytningen och remodelleringen av ECM under läkningsprocessen efter en seneskada. Vissa MMP:er, däribland MMP-1, MMP-2, MMP-8, MMP-13 och MMP-14, har kollagenasaktivitet, vilket innebär att de, till skillnad från många andra enzymer, kan bryta ner kollagen I-fibriller. MMP-1:s nedbrytning av kollagenfibrillerna tillsammans med förekomsten av denaturerat kollagen är faktorer som tros orsaka försvagning av senans ECM och en ökad risk för att en ny bristning ska inträffa. Som svar på upprepad mekanisk belastning eller skada kan cytokiner frisättas av tenocyter och kan inducera frisättning av MMP, vilket orsakar nedbrytning av ECM och leder till återkommande skador och kroniska tendinopatier.
En mängd andra molekyler är involverade i reparation och regenerering av senor. Det finns fem tillväxtfaktorer som har visat sig vara signifikant uppreglerade och aktiva under senläkning: insulinliknande tillväxtfaktor 1 (IGF-I), platelet-derived growth factor (PDGF), vascular endothelial growth factor (VEGF), basic fibroblast growth factor (bFGF) och transforming growth factor beta (TGF-β). Dessa tillväxtfaktorer har alla olika roller under läkningsprocessen. IGF-1 ökar produktionen av kollagen och proteoglykan under det första skedet av inflammationen, och PDGF finns också under de tidiga skedena efter skadan och främjar syntesen av andra tillväxtfaktorer tillsammans med syntesen av DNA och proliferationen av senceller. De tre isoformerna av TGF-β (TGF-β1, TGF-β2, TGF-β3) är kända för att spela en roll vid sårläkning och ärrbildning. VEGF är välkänt för att främja angiogenes och inducera endotelcellsproliferation och migration, och VEGF mRNA har visat sig uttryckas på platsen för seneskador tillsammans med kollagen I mRNA. Bone morphogenetic proteins (BMP) är en undergrupp av TGF-β superfamiljen som kan inducera ben- och broskbildning samt vävnadsdifferentiering, och BMP-12 specifikt har visats påverka bildning och differentiering av senvävnad och främja fibrogenes.
Effekter av aktivitet på läkning
I djurmodeller har omfattande studier utförts för att undersöka effekterna av mekanisk påfrestning i form av aktivitetsnivå på seneskada och läkning. Medan stretching kan störa läkningen under den inledande inflammatoriska fasen har det visat sig att kontrollerad rörelse av senorna efter cirka en vecka efter en akut skada kan bidra till att främja tenocyternas kollagensyntes, vilket leder till ökad draghållfasthet och diameter hos de läkta senorna och färre vidhäftningar än senor som är immobiliserade. Vid kroniska seneskador har mekanisk belastning också visat sig stimulera fibroblastproliferation och kollagensyntes tillsammans med kollagenomläggning, vilket allt främjar reparation och remodellering. För att ytterligare stödja teorin att rörelse och aktivitet hjälper till med läkning av senor har det visat sig att immobilisering av senorna efter en skada ofta har en negativ effekt på läkningen. Hos kaniner har kollagenfascikler som är immobiliserade visat minskad draghållfasthet, och immobilisering resulterar också i lägre mängder vatten, proteoglykaner och kollagenkorslänkar i senorna.
Flera mekanotransduktionsmekanismer har föreslagits som orsaker till tenocyternas svar på mekanisk kraft som gör att de kan förändra sitt genuttryck, sin proteinsyntes och sin cellfenotyp, och så småningom orsaka förändringar i senans struktur. En viktig faktor är mekanisk deformation av den extracellulära matrisen, som kan påverka aktincytoskelettet och därmed påverka cellform, motilitet och funktion. Mekaniska krafter kan överföras av fokala adhesionsställen, integriner och cell-cellförbindelser. Förändringar i aktincytoskelettet kan aktivera integriner, som förmedlar ”outside-in” och ”inside-out” signalering mellan cellen och matrisen. G-proteiner, som inducerar intracellulära signalkaskader, kan också vara viktiga, och jonkanaler aktiveras av sträckning för att låta joner som kalcium, natrium eller kalium komma in i cellen.
Samhälle och kultur
Sinka användes i stor utsträckning under hela den förindustriella eran som en tuff, hållbar fiber. Några specifika användningsområden är att använda senor som tråd för sömnad, fästa fjädrar på pilar (se fletch), surra fast verktygsblad på axlar osv. Den rekommenderas också i överlevnadsguider som ett material av vilket man kan göra starka snören för t.ex. fällor eller levande strukturer. Tendon måste behandlas på särskilda sätt för att fungera på ett användbart sätt för dessa ändamål. Inuiter och andra cirkumpolära folk använde senor som det enda snöret för alla hushållsändamål på grund av bristen på andra lämpliga fiberkällor i deras ekologiska livsmiljöer. De elastiska egenskaperna hos vissa senor användes också i sammansatta rekurvade bågar som föredrogs av stäppnomaderna i Eurasien och indianerna. Det första stenkastande artilleriet använde sig också av senornas elastiska egenskaper.
Senor är ett utmärkt material för snörning av tre anledningar: Det är extremt starkt, det innehåller naturliga lim och det krymper när det torkar, vilket gör att det inte behövs några knutar.
Kulinariska användningsområden
Senor (i synnerhet nötköttsenor) används som livsmedel i vissa asiatiska kök (ofta serveras de på yum cha- eller dim sum-restauranger). En populär maträtt är suan bao niu jin, där senan marineras i vitlök. Den finns också ibland i den vietnamesiska nudelrätten phở.
Klinisk betydelse
Skada
Senor är utsatta för många typer av skador. Det finns olika former av tendinopatier eller seneskador på grund av överanvändning. Dessa typer av skador leder i allmänhet till inflammation och degeneration eller försvagning av senorna, vilket så småningom kan leda till senruptur. Tendinopatier kan orsakas av ett antal faktorer som rör senans extracellulära matris (ECM), och deras klassificering har varit svår eftersom deras symtom och histopatologi ofta liknar varandra.
Den första kategorin tendinopatier är paratenonit, som avser inflammation i paratenon, eller paratendinalskiktet som ligger mellan senan och dess skida. Tendinosis hänvisar till icke-inflammatorisk skada på senan på cellnivå. Nedbrytningen orsakas av skador på kollagen, celler och de vaskulära komponenterna i senan och är känd för att leda till bristning. Observationer av senor som har genomgått en spontan bristning har visat förekomsten av kollagenfibriller som inte är i rätt parallell orientering eller som inte är enhetliga i längd eller diameter, tillsammans med rundade tenocyter, andra cellavvikelser och inväxning av blodkärl. Andra former av tendinos som inte har lett till bristning har också visat degeneration, desorientering och gallring av kollagenfibrillerna, tillsammans med en ökning av mängden glykosaminoglykaner mellan fibrillerna. Den tredje kategorin är paratenonit med tendinos, där kombinationer av både paratenoninflammation och senedegeneration förekommer. Den sista är tendinit, som avser degeneration med inflammation i senan samt vaskulär störning.
Tendinopatier kan orsakas av flera inneboende faktorer, bland annat ålder, kroppsvikt och näring. De extrinsika faktorerna är ofta relaterade till sport och inkluderar överdrivna krafter eller belastning, dålig träningsteknik och miljöförhållanden.
Andra djur

I vissa organismer, noterbara är fåglar och ornitschiska dinosaurier, kan delar av senan bli ossifierad. I denna process infiltrerar osteocyter senan och lägger ner ben på samma sätt som de skulle göra i sesamoidben som t.ex. patella. Hos fåglar sker förbening av senor främst i bakbenet, medan förbenade axialmuskelssenor hos ornitschiska dinosaurier bildar ett nätverk längs svansens neural- och hemalstjärna, förmodligen för att ge stöd.
Se även
| Wikimedia Commons har media som rör Senor. |
- Aponeuros
- Knälbrosk
- Chordae tendineae
- Lista över muskler i människokroppen
- Tendonskidan
- ^ Dorlands medicinska ordbok, sid 602
- ^ Caldini, E. G.; Caldini, N.; De-Pasquale, V.; Strocchi, R.; Guizzardi, S.; Ruggeri, A.; Montes, G. S. (1990). ”Distribution of elastic system fibres in the rat tail tendon and its associated sheaths”. Cells Tissues Organs. 139 (4): 341-348. doi:10.1159/000147022. PMID 1706129.
- ^ Grant, T. M.; Thompson, M. S.; Urban, J.; Yu, J. (2013). ”Elastiska fibrer är brett fördelade i senan och starkt lokaliserade runt tenocyter”. Journal of Anatomy. 222 (6): 573-579. doi:10.1111/joa.12048. PMC 3666236. PMID 23587025.
- ^ Dorlands Medical Dictionary 2012.Page 1382
- ^ a b c Jozsa, L., and Kannus, P., Human Tendons: Anatomi, fysiologi och patologi. Human Kinetics: Champaign, IL, 1997.
- ^ Lin, T. W.; Cardenas, L.; Soslowsky, L. J. (2004). ”Biomechanics of tendon injury and repair”. Journal of Biomechanics. 37 (6): 865-877. doi:10.1016/j.jbiomech.2003.11.005. PMID 15111074.
- ^ Kjær, Michael (april 2004). ”Role of Extracellular Matrix in Adaptation of Tendon and Skeletal Muscle to Mechanical Loading”. Physiological Reviews. 84 (2): 649-698. doi:10.1152/physrev.00031.2003. ISSN 0031-9333. PMID 15044685.
- ^ Taye, Nandaraj; Karoulias, Stylianos Z.; Hubmacher, Dirk (januari 2020). ”De ”andra” 15-40 procenten: The Role of Non-Collagenous Extracellular Matrix Proteins and Minor Collagens in Tendon”. Journal of Orthopaedic Research. 38 (1): 23-35. doi:10.1002/jor.24440. ISSN 0736-0266. PMC 6917864. PMID 31410892.
- ^ Fukuta, S.; Oyama, M.; Kavalkovich, K.; Fu, F. H.; Niyibizi, C. (1998). ”Identifiering av kollagen av typerna II, IX och X vid insticksstället för achillessenen hos nötkreatur”. Matrix Biology. 17 (1): 65-73. doi:10.1016/S0945-053X(98)90125-1. PMID 9628253.
- ^ Fratzl, P. (2009). ”Cellulosa och kollagen: från fibrer till vävnader”. Current Opinion in Colloid & Interface Science. 8 (1): 32-39. doi:10.1016/S1359-0294(03)00011-6.
- ^ Zhang, G. E., Y.; Chervoneva, I.; Robinson, P. S.; Beason, D. P.; Carine, E. T.; Soslowsky, L. J.; Iozzo, R. V.; Birk, D. E. (2006). ”Decorin reglerar sammansättningen av kollagenfibriller och förvärv av biomekaniska egenskaper under senans utveckling”. Journal of Cellular Biochemistry. 98 (6): 1436-1449. doi:10.1002/jcb.20776. PMID 16518859. S2CID 39384363. CS1 maint: multiple names: authors list (link)
- ^ Raspanti, M.; Congiu, T.; Guizzardi, S. (2002). ”Structural Aspects of the Extracellular Matrix of the Tendon : An Atomic Force and Scanning Electron Microscopy Study”. Archives of Histology and Cytology. 65 (1): 37-43. doi:10.1679/aohc.65.37. PMID 12002609.
- ^ Scott, J. E. O., C. R.; Hughes, E. W. (1981). ”Proteoglykan-kollagenarrangemang i utvecklande svanssenor hos råttor. En elektronmikroskopisk och biokemisk undersökning”. Biochemical Journal. 195 (3): 573-581. doi:10.1042/bj1950573. PMC 1162928. PMID 6459082. CS1 maint: multiple names: authors list (link)
- ^ Scott, J. E. (2003). ”Elasticitet i extracellulära matris ’formmoduler’ av senor, brosk osv. A sliding proteoglycan-filament model”. Journal of Physiology. 553 (2): 335-343. doi:10.1113/jphysiol.2003.050179. PMC 2343561. PMID 12923209.
- ^ McNeilly, C. M.; Banes, A. J.; Benjamin, M.; Ralphs, J. R. (1996). ”Tendon cells in vivo form a three dimensional network of cell processes linked by gap junctions”. Journal of Anatomy. 189 (Pt 3): 593-600. PMC 1167702. PMID 8982835.
- ^ ”Having a short Achilles tendon may be an athlete’s Achilles heel”. Hämtad 2007-10-26.
- ^ Young, Michael. ”A Review on Postural Realignment and its Muscular and Neural Components” (PDF).
- ^ Thorpe C.T., Birch H.L., Clegg P.D., Screen H.R.C. (2013). Den icke-kollagena matrisens roll i senans funktion. Int J ExpPathol. 94;4: 248-59.
- ^ Hulmes, D. J. S. (2002). ”Building Collagen Molecules, Fibrils, and Suprafibrillar Structures”. Journal of Structural Biology. 137 (1-2): 2-10. doi:10.1006/jsbi.2002.4450. PMID 12064927.
- ^ Silver, F. H.; Freeman, J. W.; Seehra, G. P. (2003). ”Collagen self-assembly and the development of tendon mechanical properties”. Journal of Biomechanics. 36 (10): 1529-1553. doi:10.1016/S0021-9290(03)00135-0. PMID 14499302.
- ^ Ker, R. F. (2002). ”Konsekvenserna av den anpassningsbara utmattningskvaliteten hos senorerna för deras konstruktion, reparation och funktion”. Comparative Biochemistry and Physiology A. 133 (4): 987-1000. doi:10.1016/S1095-6433(02)00171-X. PMID 12485688.
- ^ Cribb, A. M.; Scott, J.E. (1995). In Tendon response to tensile-stress – an ultrastructural investigation of collagen – proteoglycan interactions in stressed tendon,1995; Cambridge Univ Press.pp 423-428.
- ^ Screen H.R., Lee D.A., Bader D.L., Shelton J.C. (2004). ”An investigation into the effects of the hierarchical structure of tendon fascicles on micromechanical properties”. Proc Inst Mech Eng H. 218 (2): 109-119. doi:10.1243/095441104322984004. PMID 15116898. S2CID 46256718.CS1 maint: multiple names: authors list (link)
- ^ Puxkandl, R.; Zizak, I.; Paris, O.; Keckes, J.; Tesch, W.; Bernstorff, S.; Purslow, P.; Fratzl, P. (2002). ”Viskoelastiska egenskaper hos kollagen: undersökningar med synkrotronstrålning och strukturmodell”. Philosophical Transactions of the Royal Society B. 357 (1418): 191-197. doi:10.1098/rstb.2001.1033. PMC 1692933. PMID 11911776.
- ^ Gupta H.S., Seto J., Krauss S., Boesecke P.& Screen H.R.C. (2010). In situ multi-level analysis of viscoelastic deformation mechanisms in tendon collagen. J. Struct. Biol. 169(2):183-191.
- ^ Thorpe C.T; Udeze C.P; Birch H.L.; Clegg P.D.; Screen H.R.C. (2012). ”Specialisering av senans mekaniska egenskaper beror på interfascikulära skillnader”. Journal of the Royal Society Interface. 9 (76): 3108-3117. doi:10.1098/rsif.2012.0362. PMC 3479922. PMID 22764132.
- ^ Thorpe C.T.; Klemt C; Riley G.P.; Birch H.L.; Clegg P.D.; Screen H.R.C. (2013). ”Helical sub-structures in energy-storing tendons provide a possible mechanism for efficient energy storage and return”. Acta Biomater. 9 (8): 7948-56. doi:10.1016/j.actbio.2013.05.004. PMID 23669621.
- ^ Gatt R, Vella Wood M, Gatt A, Zarb F, Formosa C, Azzopardi KM, Casha A, Agius TP, Schembri-Wismayer P, Attard L, Chockalingam N, Grima JN (2015). ”Negativa Poissonkvoten i senor: Ett oväntat mekaniskt svar”. Acta Biomater. 24: 201-208. doi:10.1016/j.actbio.2015.06.018. PMID 26102335.
- ^ Batson EL, Paramour RJ, Smith TJ, Birch HL, Patterson-Kane JC, Goodship AE. (2003). Equine Vet J. |volume=35 |issue=3 |pages=314-8. Är materialegenskaperna och matrisammansättningen hos hästars böj- och sträcksenor bestämda av deras funktioner?
- ^ ScreenH.R.C., Tanner, K.E. (2012). Structure & Biomechanics of Biological Composites. In: Encyclopaedia of Composites 2nd Ed. Nicolais & Borzacchiello.Pub. John Wiley & Sons, Inc. ISBN 978-0-470-1282828-2 (pages 2928-39)
- ^ Nakagawa, Y. (1989). ”Effekten av avstängning på ultrastrukturen i akillessenen hos råttor”. European Journal of Applied Physiology. 59 (3): 239-242. doi:10.1007/bf02386194. PMID 2583169. S2CID 20626078.
- ^ Reeves, N. D. (2005). ”Influens av 90-dagars simulerad mikrogravitation på mänskliga senors mekaniska egenskaper och effekten av motåtgärder för resning”. Journal of Applied Physiology. 98 (6): 2278-2286. doi:10.1152/japplphysiol.01266.2004. hdl:11379/25397. PMID 15705722.
- ^ a b Riley, G. (2004). ”Patogenesen för tendinopati. Ett molekylärt perspektiv” (PDF). Reumatologi. 43 (2): 131-142. doi:10.1093/rheumatology/keg448. PMID 12867575.
- ^ a b c d Sharma, P. M., N. (2006). ”Biologi vid skador på senor: läkning, modellering och remodellering”. Journal of Musculoskeletal and Neuronal Interactions. 6 (2): 181-190. PMID 16849830. CS1 maint: multiple names: authors list (link)
- ^ a b c d Sharma, P.; Maffulli, N. (2005). ”Senkskador och tendinopati: läkning och reparation”. Journal of Bone and Joint Surgery. American Volume. 87A (1): 187-202. doi:10.2106/JBJS.D.01850. PMID 15634833. S2CID 1111422.
- ^ a b c d e f Wang, J. H. C. (2006). ”Mechanobiology of tendon”. Journal of Biomechanics. 39 (9): 1563-1582. doi:10.1016/j.jbiomech.2005.05.011. PMID 16000201.
- ^ Riley, G. P.; Curry, V.; DeGroot, J.; van El, B.; Verzijl, N.; Hazleman, B. L.; Bank, R. A. (2002). ”Matrix metalloproteinase activities and their relationship with collagen remodelling in tendon pathology”. Matrix Biology. 21 (2): 185-195. doi:10.1016/S0945-053X(01)00196-2. PMID 11852234.
- ^ Moulin, V.; Tam, B. Y. Y.; Castilloux, G.; Auger, F. A.; O’Connor-McCourt, M. D.; Philip, A.; Germain, L. (2001). ”Fetal and adult human skin fibroblasts display intrinsic differences in contractile capacity”. Journal of Cellular Physiology. 188 (2): 211-222. doi:10.1002/jcp.1110. PMID 11424088. S2CID 22026692.
- ^ Boyer, M. I. W., J. T.; Lou, J.; Manske, P. R.; Gelberman, R. H.; Cai, S. R. (2001). ”Kvantitativ variation i mRNA-uttrycket av vascular endothelial growth factor under tidig läkning av böjsenan: en undersökning i en hundmodell”. Journal of Orthopaedic Research. 19 (5): 869-872. doi:10.1016/S0736-0266(01)00017-1. PMID 11562135. S2CID 20903366. CS1 maint: multiple names: authors list (link)
- ^ Astrom, M.; Rausing, A. (1995). ”Chronic Achilles Tendinopathy – A survey of Surgical and Histopathologic findings”. Clinical Orthopaedics and Related Research. 316 (316): 151-164. doi:10.1097/00003086-199507000-00021. PMID 7634699. S2CID 25486134.
- ^ Berge, James C. Vanden; Storer, Robert W. (1995). ”Intratendinous ossifikation hos fåglar: A review”. Journal of Morphology. 226 (1): 47-77. doi:10.1002/jmor.1052260105. PMID 29865323. S2CID 46926646.
- ^ Organ, Chris L. (2006). ”Biomekanik hos förbenade senor hos ornithopoda dinosaurier”. Paleobiology. 32 (4): 652-665. doi:10.1666/05039.1. S2CID 86568665.